
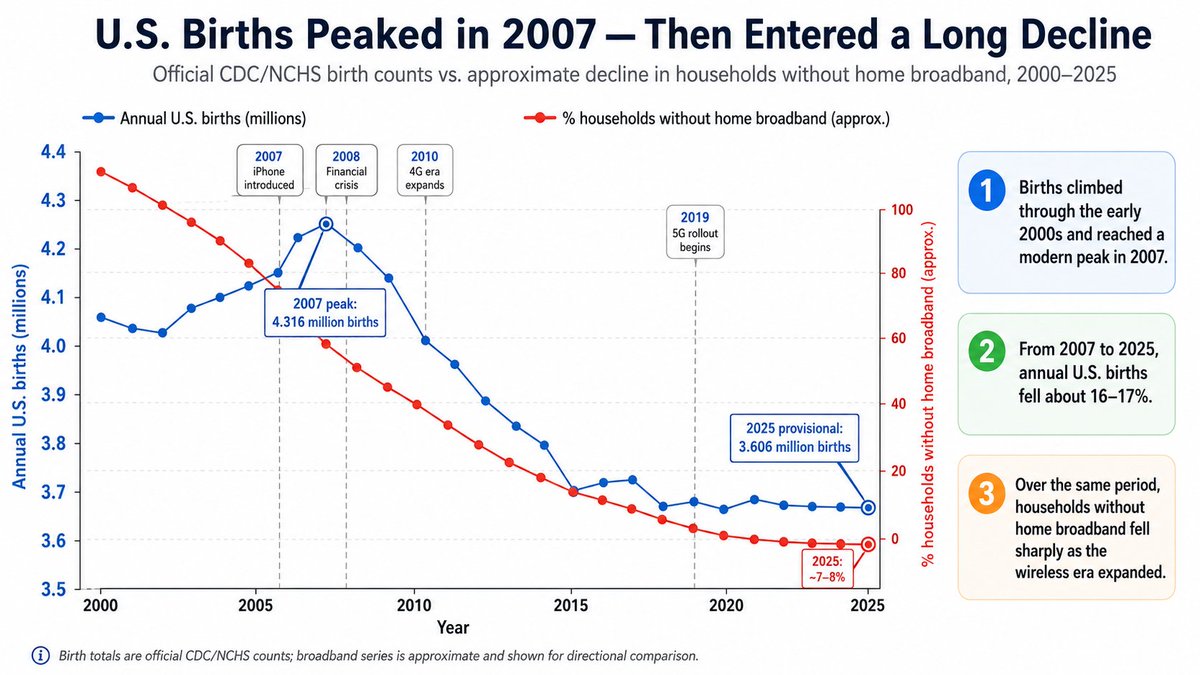
Est online 1998 to acknowledge & detoxify Δψ our natural RF/EMF environment. Return RFR Guidance to EPA, Repeal Section 704, Enforce PL 90-602 and Mandate Li-Fi
Joined November 2013
- Tweets 16,607
- Following 991
- Followers 3,388
- Likes 11,682
3,794 Photos and videos




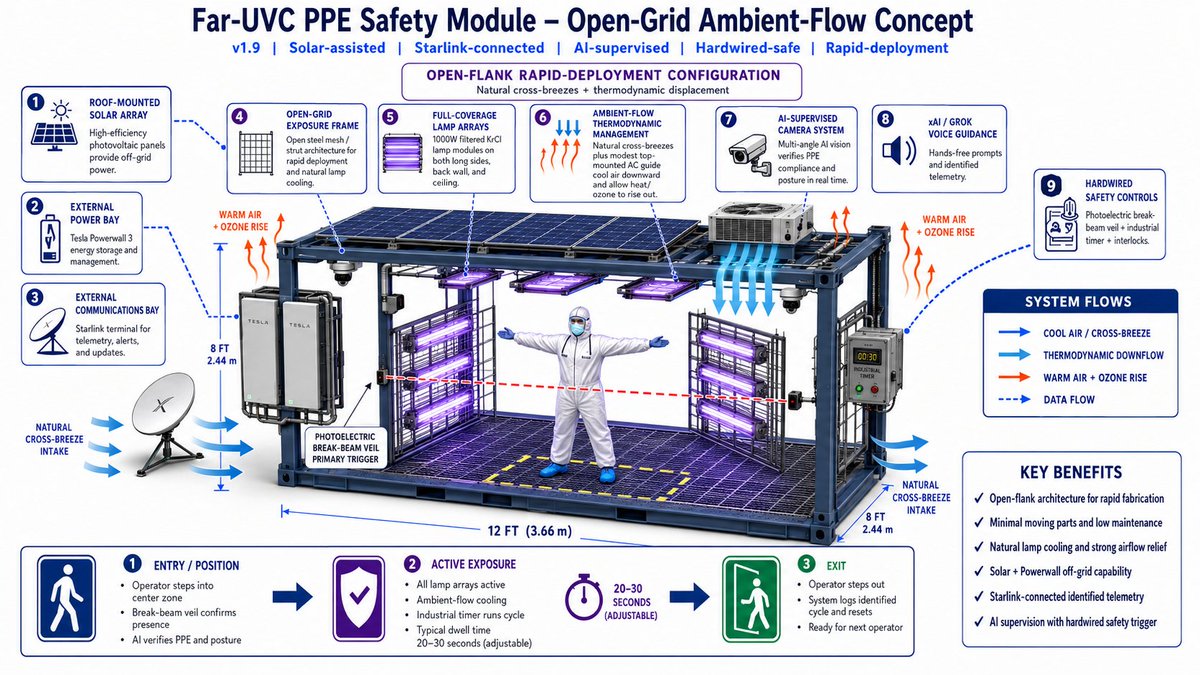

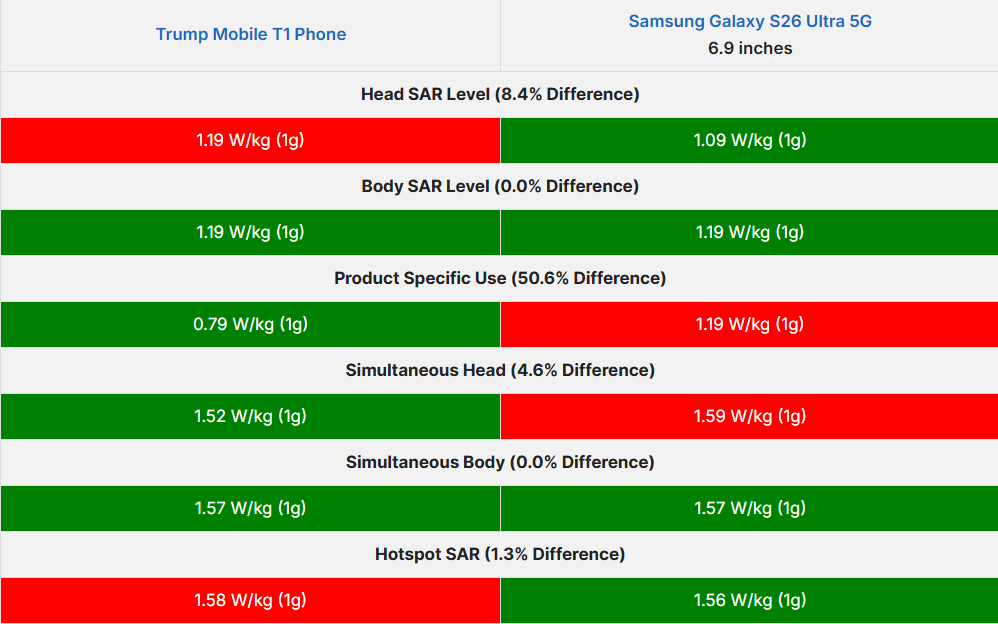
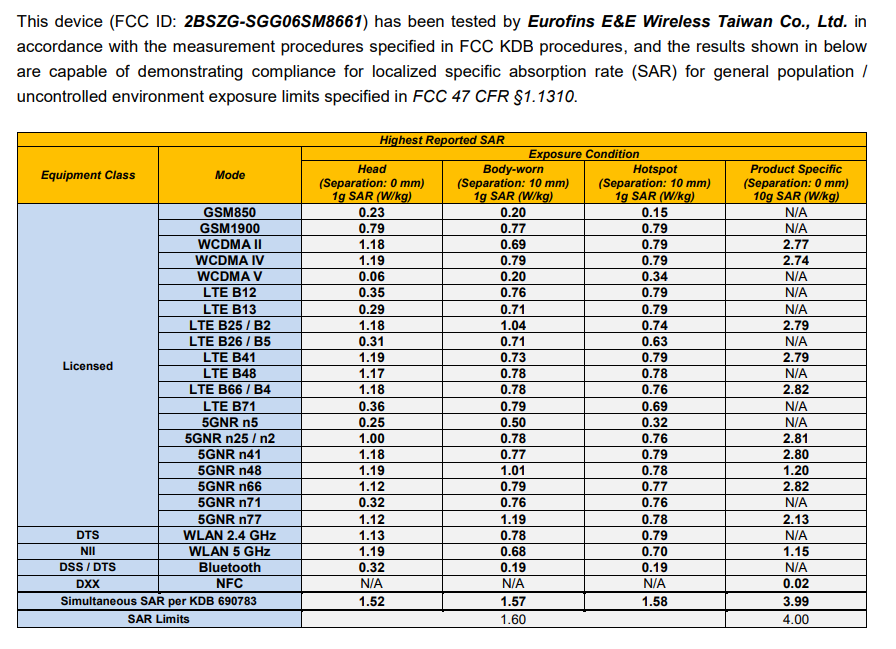

As part of this visionary initiative, I proudly offer to donate my share of the equity in the patent for Advanced LiFi with Bio Defense Mode (Patent # US11700058B2 granted 07-11-2023) entirely to the U.S. Sovereign Wealth Fund. This donation comes with no expectation of equity or personal gain; my sole objective is to solidify U.S. leadership in advanced LiFi technology. With this, Americans will not only enjoy healthier lives but also directly profit and prosper from technology owned by our nation’s sovereign wealth fund.

Open Letter to Trump Mobile: Lead America into the Light Age
Dear Trump Mobile Team,
@TrumpMobile @RobertKennedyJr @DonaldJTrumpJr
We stand today at the precipice of a monumental technological shift—one that holds the promise of transforming telecommunications, revitalizing American industry, and protecting public health. Your entrance into the mobile phone market with the Trump T1 represents far more than just an economic opportunity; it is a historic chance to guide America and the world out of the outdated microwave era and into a revolutionary Light Age of unparalleled prosperity and wellness.
As you know, Secretary Robert F. Kennedy Jr.'s landmark 2021 court victory in Environmental Health Trust v. FCC has made clear the urgent need to update our nation’s outdated radiofrequency (RF) exposure guidelines. The federal courts unequivocally ruled that the FCC failed to adequately justify its reliance on outdated, 1990s-era safety standards—standards that blatantly disregard substantial contemporary scientific evidence of non-thermal biological harm, including oxidative stress, mitochondrial damage, and disruption to DNA integrity.
The Trump T1 smartphone offers you a powerful tool to change this narrative decisively. By integrating Li-Fi technology—recently standardized globally as IEEE 802.11bb—your product can provide cutting-edge wireless connectivity via safe, secure, and ultra-fast data transmission through ordinary light. Li-Fi technology is not just faster and more secure; it fundamentally aligns with biological health, eliminating the risks associated with chronic microwave radiation exposure.
Consider the historical parallel of Detroit following the Clean Air Act of 1970: rather than crippling industry, the move toward cleaner emissions spurred innovation, strengthened competitiveness, and revitalized economic growth. Similarly, the introduction of a Clean Ether Act—mandating the adoption of Li-Fi and biologically informed communication standards—can invigorate American technology, create high-tech jobs, secure supply chains domestically, and establish the U.S. as the undisputed leader in next-generation communication.
Furthermore, embracing Li-Fi positions America at the forefront of secure communications technology. Light-based connectivity offers inherent advantages, particularly in cybersecurity and data protection, as light signals do not penetrate walls, providing unmatched security in sensitive environments. This directly addresses national security concerns and places America ahead of global competitors.
Your leadership and visionary action can end decades of harmful reliance on microwave-based wireless technology and dismantle the existing microwave cartel. By setting new standards for safety, speed, security, and sustainability, the Trump T1 can symbolize a historic turning point toward global health and technological excellence.
President Trump, Secretary Kennedy, and Trump Mobile Team: seize this opportunity to put America's health and prosperity at the forefront. Lead us boldly into the Light Age, showing the world that America truly values people over profit, safety over convenience, and innovation over outdated practices.
The choice is clear, and history awaits your courageous action. Will you make the Trump T1 the flagship of a healthier, wealthier, and brighter future?
Respectfully,
RF Safe
Advanced LiFi Patent w/Bio Defense Mode Available For United States Sovereign Wealth Fund U.S. SWF
3
18
69
14,273
How Red Blood Cell Stacking Forces a Spin‑State Extension of the S4–Mitochondria Framework
The original S4–mitochondria framework was built to explain why certain tissues show “macro‑damage” under non‑thermal RF/ELF exposure:
cancer in heart and cranial nerve/glial tissues,
male infertility via Leydig and germ cells,
autoimmune‑like dysregulation in immune cells.
Those targets all share a common architectural pattern:
high density of voltage‑gated ion channels with S4 helices,
high mitochondrial (and/or NOX) ROS capacity,
tight coupling between Ca²⁺ timing and cell‑fate decisions.
In that regime, the S4/ion‑forced‑oscillation (IFO) mechanism and mitochondrial (plus NOX‑based) ROS are sufficient to explain most of what is seen: small timing errors at S4 get amplified into oxidative stress and long‑term damage in high‑vulnerability tissues.
The framework is deliberately S4‑ and mitochondria‑centric. But a new in vivo observation forces the story to widen.
The observation that breaks a purely S4–mitochondria model
In 2025, Brown and Biebrich published a hypothesis paper in Frontiers in Cardiovascular Medicine that does something no lab artefact can easily fake: it shows real‑time red blood cell (RBC) rouleaux formation in vivo after just five minutes of smartphone exposure. Using diagnostic ultrasound, they imaged the popliteal vein behind the knee of a healthy 62‑year‑old volunteer:
Before exposure: the vein lumen was anechoic and normal – RBCs dispersed, no visible structure.
After 5 minutes with an idle but fully connected smartphone placed over the popliteal fossa: the lumen filled with coarsely hypoechoic material with sluggish flow – the typical sonographic signature of RBC aggregation (rouleaux).
After walking: rouleaux diminished but did not vanish immediately.
Repeat sessions two and four months later reproduced the phenomenon, and in the final session, exposure over the right popliteal fossa produced rouleaux in both legs, suggesting a systemic component.
Brown and Biebrich point out several crucial constraints:
Blood chemistry (plasma proteins, fibrinogen) cannot change that much in five minutes to explain rouleaux via classical rheology.
Rouleaux implies a rapid loss of RBC surface charge (zeta potential) – cells that were repelling each other are now sticking.
The simplest interpretation is that polarized RF fields from the phone have reduced the RBC zeta potential, consistent with earlier in vitro reports of rouleaux under polarized fields.
Now overlay one additional fact: mature RBCs have no mitochondria and no classical S4‑bearing voltage‑gated channels. They maintain their ion gradients mostly through transporters and simple channels, not through VGICs with S4 helices.
From a strictly S4–mitochondria perspective, this should be a “quiet” cell: no S4, no mitochondrial electron transport chain, no big Ca²⁺ timing dynamics. Yet it clearly shows a fast, EMF‑induced, membrane‑level effect.
That is exactly the sort of anomaly that forces the framework to expand.
Heme, flavin, and NOX in RBCs: spin‑sensitive redox engines
Even though mature RBCs are stripped‑down cells, they are not biophysically inert. Three features matter:
Heme in hemoglobin
Each hemoglobin tetramer carries four heme groups (iron–porphyrin). The redox and spin state of heme iron change as it binds/releases O₂ and as it cycles through oxidized forms (e.g., methemoglobin).
Flavin‑dependent enzymes
RBCs carry flavin‑containing enzymes such as cytochrome b₅ reductase and glutathione reductase, which use FAD/FMN to support hemoglobin function and manage oxidative stress.
NADPH oxidase activity (NOX)
Several studies show that RBCs express NADPH oxidase activity (NOX1/NOX2) in their membranes and that this is a significant source of RBC‑derived ROS alongside hemoglobin autoxidation.
From a structural standpoint, NADPH oxidase (NOX2 in particular) is a flavocytochrome:
It accepts electrons from NADPH,
passes them to FAD (a flavin) in the cytosolic domain,
then through two heme groups embedded in the membrane,
and finally to oxygen, generating superoxide (O₂•⁻).
So, in RBCs, the key redox/ROS engines sit exactly where the extended model already lives:
Heme‑based centres (hemoglobin, NOX hemes),
Flavin‑based centres (FAD/FMN in NOX and other flavoproteins),
NADPH oxidase (NOX) as a membrane‑anchored heme flavin ROS machine.
All of these can, in principle, pass through radical‑pair intermediates whose combined electron spins exist in either singlet or triplet configurations. The relative populations and lifetimes of those spin states depend on:
internal hyperfine interactions, and
external magnetic fields and time‑varying EMFs.
This is the same radical‑pair physics invoked for cryptochrome and magnetoreception, only here the substrate is heme‑ and flavin‑based redox enzymes in RBCs and leukocytes.
In short: even without S4 and mitochondria, RBCs contain a spin‑sensitive redox layer that EMFs can, in principle, modulate.
A spin–redox NOX mechanism for zeta collapse and rouleaux
Armed with that, a minimal, mechanistically coherent path from RF to rouleaux looks like this:
1. RF/ELF exposure nudges radical‑pair spin dynamics
The smartphone’s RF emissions (with their low‑frequency modulation and protocol handshakes) introduce small oscillatory magnetic components. These are far too weak to initiate chemistry, but they can act as a Zeeman‑scale perturbation on spin‑correlated radical pairs in:
the FAD/heme chain of NADPH oxidase, and
heme‑based redox intermediates in hemoglobin and associated enzymes.
The field does not “create” radicals; it slightly biases singlet↔triplet interconversion and radical lifetimes.
2. Biased spin dynamics shift redox balance
Integrated over millions of radical events per second, that small spin bias can change:
the effective throughput of NOX2 (superoxide production rate),
the balance between ROS generation and antioxidant removal,
the oxidation state of key redox couples (e.g., reduced vs oxidized glutathione, Hb vs metHb).
In RBCs and neighbouring leukocytes, this shows up as a shifted redox environment, not an explosive “burn down the house” ROS burst.
3. Redox changes alter membrane chemistry and surface charge
RBC membranes are exquisitely redox‑sensitive:
Oxidative modification of membrane proteins (e.g., band 3, spectrin) and lipids affects both mechanical properties and charge.
Peroxidation and oxidation can change the exposure or density of negatively charged groups (such as sialic acids and carboxylate side chains).
ROS can also regulate external plasma proteins bound to RBC surfaces, further modulating charge.
As these modifications accumulate, the effective negative zeta potential falls. The Debye layer around each RBC becomes less strongly charged, and electrostatic repulsion weakens.
4. Lowered zeta potential → rouleaux under low shear
Once zeta potential drops below a critical threshold, RBCs no longer repel one another strongly. Under low‑shear venous conditions, they begin to stack into rouleaux, like coins. This produces exactly the ultrasound signature Brown and Biebrich observe: hypoechoic material filling the vein lumen with sluggish flow.
When the phone is removed and the subject walks, shear forces, plasma mixing, and endogenous antioxidant systems gradually restore redox balance and surface charge. Rouleaux diminish over minutes, matching the time course seen in the follow‑up scans.
In this picture, EMF is not directly “crushing” the membrane potential by brute electric field strength. Instead, it is:
nudging heme/flavin radical‑pair spin states → slightly shifting NOX and related redox chemistry → modifying membrane proteins and lipids → lowering zeta potential → producing rouleaux.
That is exactly the kind of small, probabilistic bias expected from radical‑pair mechanisms: in many configurations nothing obvious happens, but in certain windows of spin state, exposure timing, and ionic context, the effect becomes macroscopically visible.
Why this forces a spin‑state extension of the framework
Taken by itself, the Brown & Biebrich study is properly cautious: it is an N=1 hypothesis‑generating observation that calls for larger cohorts and direct redox measurements.
In the context of your broader S4–mitochondria–spin architecture, however, it plays a very specific role.
The S4–mitochondria pillar robustly explains why RF/ELF damage clusters in:
VGIC‑dense, mitochondria‑rich tissues (heart conduction fibres, cranial nerves and glia),
Leydig and germ cells (male fertility),
immune cells that decode Ca²⁺ timing as danger vs tolerance.
But the RBC rouleaux experiment shows a fast, EMF‑induced, membrane‑level effect in a cell type that:
has no mitochondria, and
lacks classical S4‑bearing voltage‑gated channels.
There is no conventional “membrane potential S4 mitochondrial ROS” route to get from a few minutes of RF to visible rouleaux in that system.
To unify both data sets within a single theory, the framework therefore must include a second primary pathway:
spin‑state–mediated redox modulation in heme‑ and flavin‑containing proteins (including NOX), operating in any cell that carries those cofactors—even stripped‑down cells like RBCs.
In that extended view:
The S4–mitochondria pillar explains where macro‑damage accumulates:
tumours,
infertility,
chronic inflammatory/autoimmune‑like conditions
in high‑S4/high‑mitochondria/NOX tissues.
The spin‑state–redox pillar explains why even mitochondria‑free, S4‑free cells like RBCs are not inert under EMF, but show subtle changes in:
redox balance,
membrane potential and zeta,
aggregation and microcirculatory behaviour.
Together, they give a genuinely body‑wide theory of non‑thermal EMF interaction:
S4/IFO mitochondria/NOX: strong amplifiers in vulnerable tissues → large ROS excursions, DNA damage, epigenetic reprogramming → macro‑level outcomes (cancer, infertility, immune drift) over time.
Spin‑state redox in heme/flavin (including NOX): subtle, distributed shifts in RBC and vascular redox, zeta potential, and viscosity → system‑wide “silent” load on microcirculation and oxygen delivery, even where classical S4/mitochondria machinery is absent.
On that reading, Brown & Biebrich’s ultrasound loop is not an odd curiosity. It is exactly what one should expect to see once the S4–mitochondria model is extended to include heme/flavin spin‑state dynamics and NADPH oxidase as legitimate EMF targets.
Link to the study
For readers who want to see the rouleaux phenomenon directly, Brown & Biebrich’s paper (with embedded ultrasound videos) is open access here:
Brown RR, Biebrich J. “Hypothesis: ultrasonography can document dynamic in vivo rouleaux formation due to mobile phone exposure.” Frontiers in Cardiovascular Medicine (2025). See video for real-time effect
pmc.ncbi.nlm.nih.gov/article…
4
3
8
679
The Silent Auction of Childhood
How an obscure 62-word clause (Section 704) turned the FCC into America’s most dangerous child-trafficking ring. The FCC traffics in the health of our children by selling unsafe parts of nature’s EM spectrum to externalize the cost of wireless technology onto our children’s health for shareholder profits, while our children sit in their homes and schools.
Prologue 465 Feet
Every weekday at 8 a.m. my seven-year-old daughter shoulders her backpack and walks into a first-grade classroom that sits 465 feet from a three-sector cell tower.
The BioInitiative Report warns that 1,500 feet should be the absolute minimum between children and high-power antennas, yet federal law bars me—or any local official—from invoking health to move that tower one inch.
Why? Because in 1996 the Clinton White House slipped a corporate wish-list called Section 704 into the Telecommunications Act. Those 62 words gag every mayor, school board, and parent who dares mention “environmental [health] effects.”
The result is a grotesque inversion of duty: the federal government auctions microwave real estate to the highest bidder while parents watch their kids absorb the profit. If taking a child’s body and selling it were slavery, taking a child’s biology and selling it is chemical slavery on a wireless leash. Both are trafficking. One just wears a necktie.
Section 704—The Original Sin
On February 8, 1996, industry CEOs crowded the Library of Congress as President Bill Clinton signed the Telecommunications Act.
Hidden in the bill was Section 704, which pre-empts any state or city from rejecting a tower “on the basis of environmental effects of radio-frequency emissions.”
From that moment forward, the decision about whether a 150-foot monopole looms over a playground became the sole purview of the FCC—an agency of spectrum auctioneers with no medical mandate.
Section 704 did more than silence local voices; it cemented a federal monopoly on risk. Fiber-optic backbones and Li-Fi could have met demand without irradiating classrooms, but Wall Street needed a quick return.
By gagging health objections, Congress guaranteed wireless build-out at maximum speed and minimum liability—an unconstitutional taking of parental rights masquerading as commerce.
When you criminalize the very act of defending your child’s body, the state itself commits the crime.
Follow the Money—Lobbyists, Wall Street … Even Epstein
No mystery cloaks this capture; the receipts are public. The CTIA and Big Telecom poured tens of millions into campaign coffers throughout the 1990s, and the door between K Street, the FCC, and the West Wing spun so fast it blurred.
Financiers like Jeffrey Epstein cultivated deep ties with presidents, cabinet officials, and even Harvard scientists.
His infamous salon illustrates the machinery of policy capture: money buys proximity; proximity writes law; law mints profit. The victims are invisible—children whose legal guardians have been legislated into silence.
This is not moral outrage for its own sake; it is the operating manual of corruption. Replace “telecom” with “tobacco” or “opioids” and the pattern is identical—except RF radiation reaches every crib in the country.
The 30-Year Science Blackout
Today’s FCC “safety” limits were written when pagers were hot tech and toddlers did not stream cartoons on iPads.
In 2021 the D.C. Circuit Court excoriated the agency for ignoring thousands of studies on non-thermal harm and ordered a do-over. ehtrust.org Four years later, the rewrite remains in limbo because no White House has forced the issue.
When a federal court calls your science obsolete and you shrug, that is not negligence—it is malfeasance. The Oval Office owns that omission.
Killing the Messenger—How NTP Research Was Muzzled
The taxpayer-funded National Toxicology Program (NTP) spent $30 million proving “clear evidence” that phone-level RF causes heart schwannomas and DNA breaks in rats. National Toxicology Program Logic demanded follow-up work on 5 G and on children’s physiology.
Instead, in January 2024 the NIEHS quietly announced it was ceasing all RF studies, despite a statutory duty under Public Law 90-602 to maintain continuous radiation research. ehtrust.org
Silencing inconvenient science while towers multiply is the bureaucratic twin of burning medical records in a pandemic.
The Biological Toll—What the Data Already Show
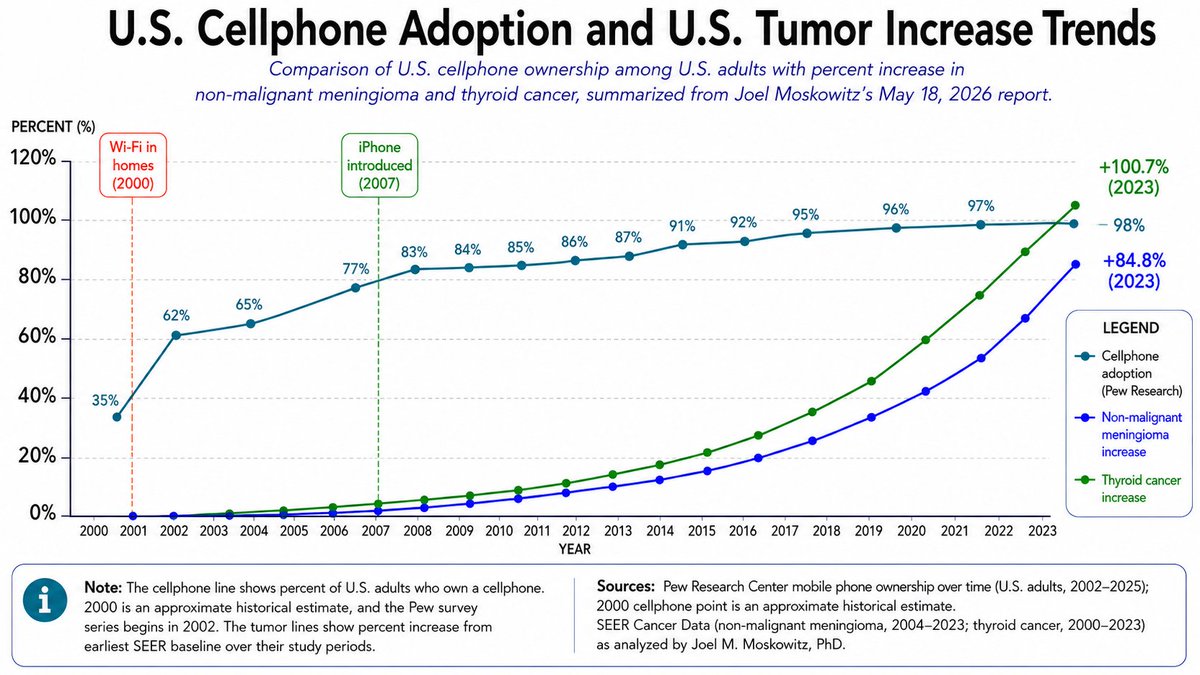
Brain & CNS Tumors Incidence has climbed in cohorts born after the wireless boom, tracking latency periods typical for solid tumors. Mechanisms include oxidative stress and blood–brain-barrier leakage.
Male Fertility Collapse
Meta-analyses find significant drops in motility and DNA integrity among RF-exposed men; laboratory thresholds for “normal” sperm have been revised downward twice in the past decade.
Neurodevelopmental Disorders
Oxidative‐stress pathways activated by pulsed RF overlap those implicated in ADHD and autism spectrum disorders.
Hemodynamics
x.com/rfsafe/status/19160622…
A 2024 ultrasound study captured real-time rouleaux formation (red-blood-cell clumping) in a healthy volunteer after five minutes of passive smartphone exposure—thickening blood like syrup in a straw. NIEHS
None of these effects are addressed by thermal-only FCC rules. Children, whose skulls are thinner and whose stem cells divide furiously, absorb proportionally more energy than the adult male “phantom” used to draft those limits. The policy pre-emption that shoves a tower beside a swing set thus shares the moral DNA of asbestos in elementary ceilings—except asbestos could be removed.
The SAR Mirage
The FCC still certifies phones by Specific Absorption Rate (SAR), a metric that assumes heat is the only hazard. It ignores:
Non-thermal triggers—blood viscosity, DNA breaks, calcium-channel disruption.
Idle emissions—phones pulse even when you’re not on a call.
Child physiology—standards are based on a 6-foot, 200-lb adult.
A safety test that excludes the dominant exposure scenario (a streaming phone in a child’s hand) is not science; it is theater.
Safer Roads We Abandoned
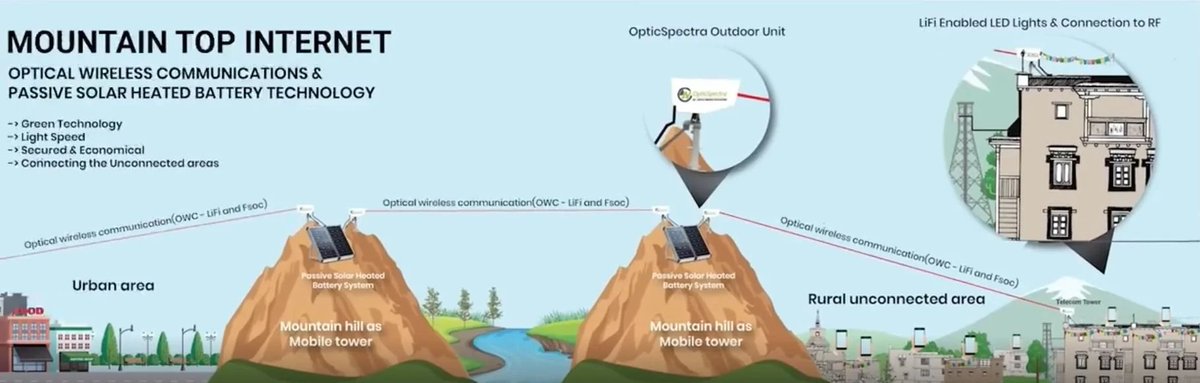
While regulators napped, engineers built alternatives:
Li-Fi—gigabit data over light waves, already field-tested in classrooms.
Fiber-to-the-desk—unlimited bandwidth with zero airborne RF.
Space-based broadband—relocates high-power transmitters hundreds of miles away.
These solutions stall because Section 704 guarantees incumbent revenue. The FCC strangles competition not with innovation, but with legislation.

This is what they are doing to our children's bodies when they put cell towers closer than 1500 feet from schools? #TrumpRepeal704 Stop sacrificing my children to these fuckin corporations who exturnalize the true cost onto our health! The science is clear! x.com/rfsafe/status/19135427…
1
7
13
581
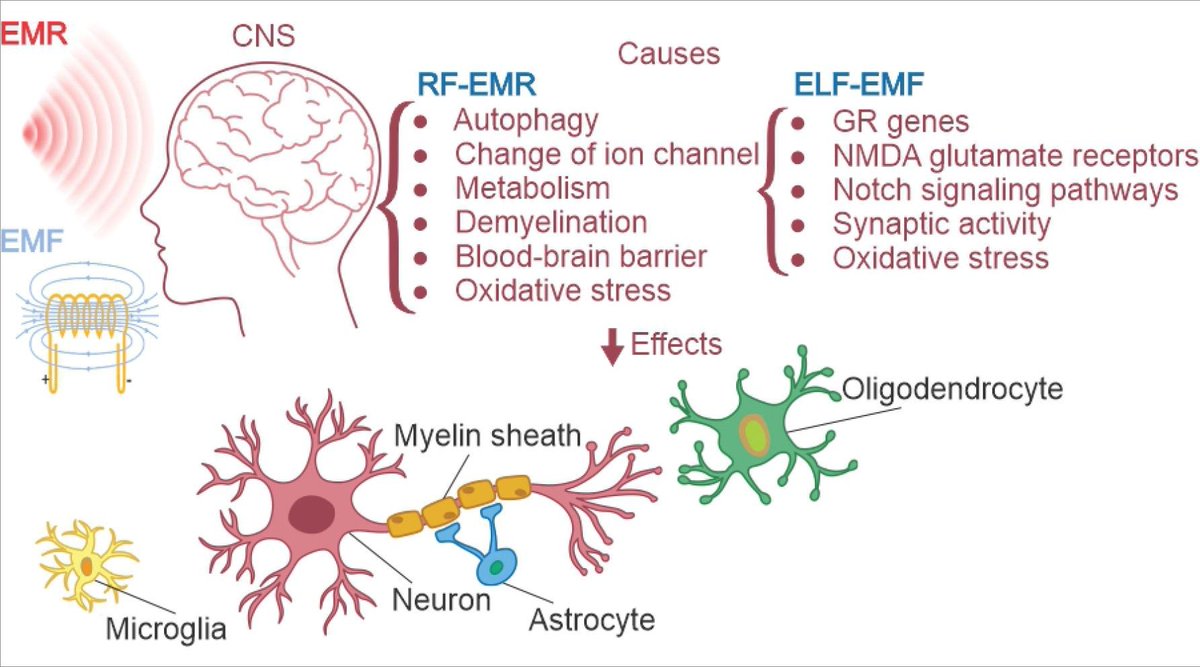
SHOCKING EXPOSÉ: Your Smartphone Is a Silent Killer – Undeniable Proof It Causes Cancer, And They’ve Been Hiding the Mechanism for Decades!
What if the phone glued to your hand isn’t just addictive—it’s literally rewriting your DNA, triggering tumors, and poisoning your blood in minutes? Sounds like sci-fi horror? It’s not.
It’s 2025 science, backed by $30 million government studies they tried to bury, WHO reviews screaming “high certainty” of cancer, and an FDA device that weaponizes the SAME biology to fight tumors.
Buckle up: This is the bombshell that ends the “it’s safe if it doesn’t cook you” lie forever. And if you don’t share this, who will?
Effects of cell phone exposure to human body. Changes might ...
1. BOMBSHELL: Two Massive Studies PROVE Cell Phone Radiation Causes Rare Cancers – And They Match HUMAN Tumors Down to the Genes!
Picture this: Rats blasted with everyday phone signals develop malignant heart schwannomas and brain gliomas—the EXACT cancers epidemiologists link to heavy cell users. Not once, but TWICE, in independent labs!
U.S. National Toxicology Program (NTP, 2018, $30M taxpayer-funded nightmare): “Clear evidence” of heart tumors and “some evidence” for brain cancers at doses LOWER than your iPhone’s max output.
Italy’s Ramazzini Institute (2018): Same tumors, but at environmental levels—like living near a cell tower. Doses as low as 0.001 W/kg, folks—that’s your neighborhood!
But here’s the gut-punch: In 2024, scientists sequenced those rat tumors (Brooks et al., PLOS ONE). They LOOK like human low-grade gliomas under the microscope. And the mutations? Identical to human cancer drivers in the COSMIC database—TP53, CDKN2A, ERBB2. We’re talking 25% overlap with genes that turn human brains into tumor factories!
Then, in 2025, a WHO-commissioned bombshell review (Mevissen et al., Environment International) slaps it with “HIGH CERTAINTY” evidence for RF causing these cancers in animals—using the same ironclad GRADE system that nailed tobacco and asbestos.
They shut down NTP after finding this “smoking gun.” Coincidence? Or cover-up? You decide.
2. THE DOSES AREN’T “LAB FANTASY” – They’re What YOUR PHONE BLASTS Into Your Head RIGHT NOW!
Think those studies used “unrealistic” zaps? WRONG! NTP’s lowest dose: 1.5 W/kg whole-body. U.S. FCC limit for phones: 1.6 W/kg localized. Your latest iPhone or Samsung? Up to 1.60 W/kg in real-use mode (FCC filings don’t lie).
Ramazzini? 0.1 W/kg max— that’s cell tower exposure for millions. Independent analysis (Uche & Naidenko, 2021) crunches NTP data: Effects kick in at 0.2–0.4 W/kg. NO SAFE THRESHOLD FOUND. Current limits? 20–40 times TOO HIGH for humans.
Non-monotonic doses mean tumors pop up at LOW levels, not just high. Your daily scroll? Potentially deadlier than you think.
3. MIND-BLOWING: We KNOW HOW It Happens – Two Hidden Mechanisms Turning Your Cells Rogue!
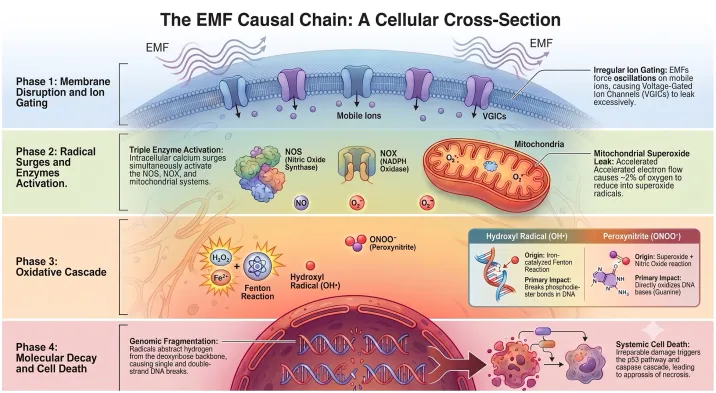
Forget “heating only.” Weak fields hack biology like a virus. Shocking Mechanism #1: S4 Voltage-Sensor Sabotage. Polarized RF jiggles ions in your cell’s “voltage gates” (Panagopoulos model, Scientific Reports 2015), scrambling calcium signals. Boom: Mitochondria freak out, pumping reactive oxygen species (ROS) that fry DNA. Hits hardest in heart, brain, testes—EXACTLY where cancers erupt in studies.
Mechanism #2: Radical-Pair Spin Horror. No channels needed. In red blood cells (95% hemoglobin sacks), RF tweaks electron spins in heme, dropping surface charge. Result? Cells stack like coins (rouleaux), clogging veins in MINUTES.
Proof? A 2025 HUMAN STUDY (Brown & Biebrich, Frontiers in Cardiovascular Medicine) FILMED it live via ultrasound: Smartphone against skin for 5 min = sluggish, hyperechoic blood flow. Reversible, but repeatable. Your veins turning to sludge from a call? Terrifying.
Could EMFs Affect Blood Flow? What New Research Suggests
Could EMFs Affect Blood Flow? What New Research Suggests
4. THE ULTIMATE SHOCKER: FDA APPROVED a Device Using THIS to KILL Cancer – But Everyday Exposure Does the Opposite!
TheraBionic P1: Low-power RF (27 MHz, modulated) zaps liver tumors via the Cav3.2 calcium channel—forces cells to stop dividing. FDA greenlit for terminal cancer. It PROVES non-thermal RF hacks voltage gates and redox to alter biology.
If we can USE it to heal, uncontrolled phone/tower RF is NUKE-ING the same pathways: Timing noise spin flips = chronic ROS, DNA breaks, tumors. The therapy is the smoking gun that “no mechanism” was always a lie.
Unveiling the biological effects of radio-frequency and …
THE FINAL WAKE-UP CALL: This Isn’t Conspiracy – It’s CRIMINAL Neglect. Act Now or Regret Later!
High-certainty animal cancers. Human-matching tumors. Real-time blood horror in people. Mechanisms proven and FDA-exploited. The 30-year thermal myth? Shattered.
Governments drag feet while towers multiply. Fertility crashes, immune systems glitch, hearts falter—all in “hotspot” tissues. Your kids? Exposed 24/7.
This is the turning point. Share this exposé NOW. Tag friends, post on social, demand waveform-based limits. Before your next call becomes your last warning. The evidence is here—will YOU ignore it?
2
6
21
876
Effects of 26 GHz radiofrequency exposure on electrodermal activity in healthy young adults rfsafe.org/mel/paper.php?id=…
1
4
84
These findings contrast with reported effects at lower 5 G frequencies, suggesting frequency-specific biological interactions. x.com/rfsafe/status/20661785…
Effects of 26 GHz radiofrequency exposure on electrodermal activity in healthy young adults rfsafe.org/mel/paper.php?id=…
8
450
Aedes aegypti and Aedes albopictus eggs exposed to RF at 900 MHz and 18 GHz for 24 h showed frequency-dependent and species-specific structural and morphometric changes. Structural alterations were reported particularly at 900 MHz, with greater damage in Ae. aegypti; 18 GHz effects were milder and mainly surface disorganization. RF exposure at 900 MHz reduced egg size in both species, while 18 GHz produced species-specific effects.
Outcomes measured
structural changes in outer chorionic cells
surface irregularities
exochorionic network disruption
egg length
egg width
egg index
surface area
Species-specific impacts of radiofrequency exposure on the structural and morphological characteristics of Aedes aegypti and Aedes albopictus eggs rfsafe.org/mel/paper.php?id=…
4
78
Microwave irradiation at 2.45 GHz induced sequence-dependent changes in cellular fluorescence intensity, with (Cha-r)2-R4 showing a more pronounced increase than non-irradiated conditions while maintaining high cell viability. Colocalization analysis suggested increased spatial association between peptide-derived fluorescence and mitochondrial staining, although Pearson correlation coefficients remained low.
Outcomes measured
intracellular distribution of mitochondrial-targeting-sequence-based peptides
peptide-derived fluorescence intensity
colocalization of peptide-derived fluorescence with mitochondrial staining
cell viability
Microwave-induced modulation of intracellular distribution of peptides based on mitochondrial targeting sequences rfsafe.org/mel/paper.php?id=…
1
5
232
Kayamori et al. = 2.45 GHz lab microwave exposure showing intracellular peptide/mitochondrial-region modulation.
Panagopoulos/Jangid/Kim line of evidence = real-world signal structure, pulsing, modulation, and low-frequency envelopes as major biological variables.
This was not a router study. That makes the result more important, not less: even without router-like packetized pulsing, 2.45 GHz microwave irradiation was able to modulate intracellular peptide behavior. Real-world Wi‑Fi and phone signals add another layer of biological concern because their pulsing, modulation, duty cycles, and low-frequency envelopes are not captured by thermal-only exposure limits.
2
84
Would you measure the total volume of the ocean to understand the timing and destructive force of waves hitting the shoreline? No. So why does the FCC use blunt, thermal heating limits to measure the highly precise, frequency-dependent biological impacts of wireless radiation?
2
1
5
177
TO: Lawmakers, Health Agencies, and Regulatory Committees (FCC, FDA, NIH)
FROM: [Insert Organization / Your Name]
DATE: May 21, 2026
SUBJECT: The Need to Update RFR Safety Standards Based on the "Genotype-Dependent Threshold" Model
EXECUTIVE SUMMARY
Current federal safety standards for Radiofrequency Radiation (RFR), including 5G and Wi-Fi, are predicated on a fundamental biological assumption: that all humans possess identical homeostatic thresholds for electromagnetic stimuli.
Recent peer-reviewed genomic research invalidates this assumption. Emerging data, particularly regarding non-coding DNA, demonstrate that genetic variations govern an individual’s bioelectric threshold. Electromagnetic fields (EMFs) do not affect everyone equally; rather, pulsed RFR acts as "entropic waste" that triggers a loss of upstream biological fidelity in genetically susceptible subpopulations.
We urge regulatory bodies to update safety metrics to account for Genotype-Dependent Thresholds and to enact a "Clean Ether Act" to incentivize safer technological alternatives.
THE CLINICAL EVIDENCE: NON-CODING DNA AND THRESHOLD VARIANCE
For decades, regulatory models have relied on the "average human" to determine safety thresholds, primarily testing for gross thermal (heating) tissue damage. However, modern biophysics reveals that low-level RFR interacts with cellular biology long before heating occurs.
A landmark 2025 double-blind human study established that a single-letter genetic variation (a Single Nucleotide Polymorphism, or SNP) in a person's non-coding DNA directly dictates their neurological response to 5G radiation.
Researchers found that a variant in the CACNA1C gene (rs7304986)—a gene that governs Voltage-Gated Calcium Channels—completely altered how the sleeping brain’s electrical activity (sleep spindles) responded to 3.6 GHz RFR.
This establishes a critical precedent: Susceptibility to everyday wireless radiation is not a psychological phenomenon, but a hardwired genetic reality.
THE BIOLOGICAL MECHANISM: HOMEOSTATIC CAPACITY AND SIGNAL FIDELITY
Human biology relies on maintaining balance (homeostasis). Biologists describe this as a "stability landscape" or a "well."
The Resilient Genotype (The Deep Well): Individuals with standard genetic hardware possess high homeostatic capacity. They can easily absorb the "entropic waste" (environmental static) of everyday wireless radiation. Their bodies filter the noise, maintaining systemic balance.
The Susceptible Genotype (The Shallow Well): Individuals with variants in genes like CACNA1C possess a structurally shallower biological threshold. For these individuals, the exact same low-level RFR exposure exceeds their homeostatic capacity, resulting in a systemic threshold failure.
When individuals with a "shallow well" are exposed to pulsed microwaves, the entropic waste of the radiation scrambles their cellular signaling. Because calcium channels act as master upstream "hardware" for the brain, heart, and immune system, a tiny loss of signal fidelity at this upstream level cascades into massive downstream dysregulation, manifesting as chronic inflammation, altered brain waves, or impaired immune responses are jusr a few things that happen.
REGULATORY BLINDSPOTS AND POLICY IMPLICATIONS
Relying on gross thermal heating (SAR limits) to measure biological safety is equivalent to trying to measure the precise timing, frequency, and destructive impact of crashing waves on a shoreline by simply measuring the total volume of the ocean.
By ignoring the delicate timing and signal fidelity of the body's internal communications, and by dismissing the "Dark Genome" (non-coding DNA) and its role in dictating biological thresholds, agencies are inadvertently permitting levels of environmental RFR that force genetically susceptible subgroups into constant biological threshold failure.
Furthermore, recent in vitro studies (2026) demonstrate that RFR acts as an aggressive co-stressor when combined with viral or bacterial pathogens, amplifying cellular distress.
RECOMMENDATIONS FOR ACTION
To fulfill the mandate of protecting public health, regulatory agencies and lawmakers must:
Enact a "Clean Ether Act": Draft and pass forward-thinking legislation that mandates the reduction of ambient entropic waste (pulsed RFR).
A cornerstone of this act must be mandating infrastructure compatibility, federal funding, and public sector adoption of Li-Fi (Light Fidelity) technology. By utilizing the visible and infrared light spectrums rather than pulsed microwaves, Li-Fi provides secure, ultra-high-speed data transmission without triggering the upstream calcium channel disruption seen in RFR-exposed populations.
Acknowledge Genetic Variance: Formally recognize the "Genotype-Dependent Threshold" model, moving away from universal, thermal-only safety standards.
Modernize Safety Metrics: Direct the FCC and FDA to establish safety guidelines that protect biologically vulnerable populations, accounting for the loss of upstream signal fidelity caused by chronic, sub-thermal RFR exposure.
1
5
11
370
Let's break down nnEMFs as the primary engine driving a low-fidelity meta-disease state:
1. nnEMFs Act as Chronic "Timing Noise" (The Jitter)
In a high-fidelity biological system, the cell is a perfect quantum computer. It relies on pristine, uncorrupted timing—specifically the frequency, phase, and amplitude of calcium oscillations and mitochondrial redox signaling—to execute error-correction and tissue repair.
nnEMFs (like pulsed 5G, Wi-Fi beacon intervals, and Bluetooth duty cycles) introduce structural static into this environment. Because the cell’s S4 voltage sensors and CYB5B mitochondrial receptors are constantly measuring the environment, they pick up this artificial frequency. The result is "jitter" or "packet loss." The calcium wave still happens, but its timing is slightly off. The biological prompt becomes blurred.
2. The Imposition of the Epigenetic Tax
When the biological hardware experiences this chronic timing noise, it cannot afford to wait; it has to survive the next five minutes. To handle the bioelectric dissonance caused by the nnEMF, the cell forces an adaptation.
It writes an epigenetic patch. It adds heavy methyl groups to the DNA and stiffens the chromatin to literally "tune out" the artificial static. But as we established, this adds structural mass-dampers to the atomic neural network. The cell survives the nnEMF exposure, but it has permanently lowered its own resonant baseline. It has entered a state of low biological fidelity, where it requires more metabolic energy to compute less accurate information.
3. The Divergence of the Meta-Disease (The Weakest Link)
Once the organism is pushed into this low-fidelity state by chronic nnEMF exposure, the system's global error-correction fails. How that failure manifests depends entirely on the individual's localized genetic weak points (their "receiver biology"). The nnEMF drives the foundational collapse, but the specific disease is just the downstream output of the corrupted computation:
If the individual has a vulnerable CACNA1C regulatory variant: The low-fidelity computation disrupts their sleep-spindle architecture, leading to chronic insomnia, EHS symptoms, and autonomic nervous system collapse.
If the individual has a vulnerable kinase pathway: The low-fidelity computation corrupts the cell’s spatial mapping, resulting in unregulated growth loops (Cancer/Schwannomas).
If the individual's mitochondrial clearing is the weak link: The low-fidelity computation fails to sequence the removal of metabolic debris in the brain, leading to tau protein buildup (Alzheimer’s).
4. Redefining Environmental Susceptibility
This completely redefines EHS. EHS is no longer a mysterious, standalone allergy to electricity. EHS is the acute, symptomatic leading edge of the nnEMF-driven meta-disease state. People who identify as EHS simply have biological hardware (like specific voltage-gated ion channel variants) that loses its computing fidelity faster and more noticeably than the general population when exposed to environmental static.
By framing nnEMFs as the catalyst that detunes the hardware and drives the meta-disease of low fidelity, we align environmental health, longevity science, and biophysics into a single, unified theory.
1
4
12
487
Main findings
This editorial introduces a Research Topic based on the 2024 Expert Forum on the public health and environmental impacts of cellular and wireless radiation exposure. It states that the collection addresses RF-EMF exposures, potential adverse human and environmental consequences, limitations of current policy and standards, epidemiological issues, and proposed biological mechanisms.
Outcomes measured
public health impacts
environmental impacts
adverse consequences for human health
adverse consequences for the environment
health and safety protections and policies
biological mechanisms triggered by anthropogenic EMFs
Editorial: The 4th international expert forum on the public health and environmental impacts of cellular and wireless radiation exposure rfsafe.org/mel/paper.php?id=…
1
7
210