Ashary Nurdin retweeted

‼️🚨Russian Defense Ministry is investigating 240 disease causing pathogens discovered in US-sponsored biolabs in Ukraine including anthrax and cholera amongst others…
Pathogens are studied and developed with intent of use for "offensive actions."
97
2,526
5,333
103,293
Unhinged retweeted

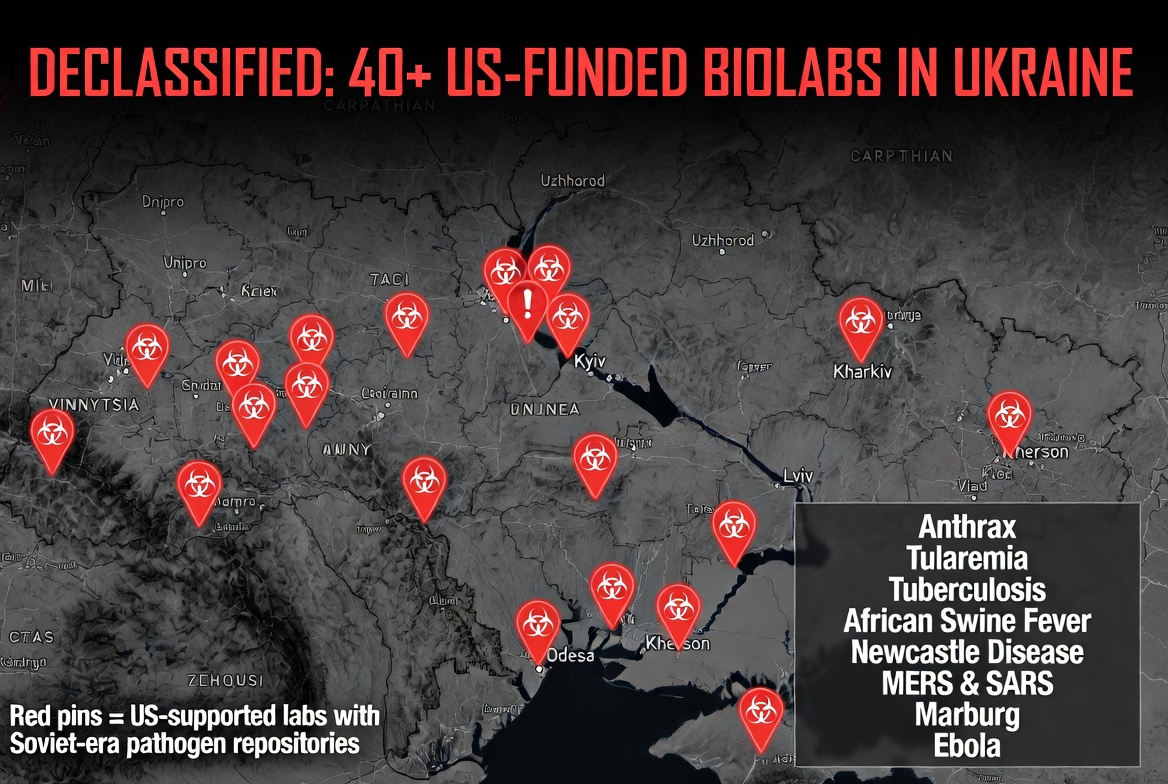
40 US-FUNDED LABS IN UKRAINE STOCKED WITH DEADLIEST PATHOGENS
Declassified documents confirm over 40 US-funded biolabs across Ukraine stored bioweapons including:
- Anthrax
- Tularemia
- Tuberculosis
- African Swine Fever
- Newcastle Disease
- MERS & SARS
- Marburg
- Ebola
These labs were part of networks directly tied to US entities:
- Black & Veatch
- CDC
- Metabiota
- EcoHealth Alliance
- Battelle
- CH2M Hill
- Ukrainian Ministry of Health US DoD contractors
Maps in the docs show the sprawling network. This wasn’t “health research” — this was a massive biolab empire on Russia’s border, funded by American taxpayers.
Who authorized this? Why were these deadly pathogens there? And why did the media and officials deny it for years while calling it “Russian disinformation”?
This needs full Congressional investigation NOW. Heads must roll.
America deserves answers. The world deserves the truth.
Juan O Savin

5
84
101
2,069
TheKnightofEvening retweeted

Key human defenses against pathogens were forged billions of years ago in microbial battles with viruses.
Learn more: scim.ag/49pOrM8 @NewsfromScience

1
3
9
1,266
Sophie retweeted

Jun 13
CONFIRMED: Barack Obama Was Involved in Creation of US Biolabs Handling Especially Dangerous Pathogens in Ukraine
Link in the comment section:

361
5,853
10,737
165,905
Annaemaila retweeted

40 US-FUNDED LABS IN UKRAINE STOCKED WITH DEADLIEST PATHOGENS
Declassified documents confirm over 40 US-funded biolabs across Ukraine stored bioweapons including:
- Anthrax
- Tularemia
- Tuberculosis
- African Swine Fever
- Newcastle Disease
- MERS & SARS
- Marburg
- Ebola
These labs were part of networks directly tied to US entities:
- Black & Veatch
- CDC
- Metabiota
- EcoHealth Alliance
- Battelle
- CH2M Hill
- Ukrainian Ministry of Health US DoD contractors
Maps in the docs show the sprawling network. This wasn’t “health research” — this was a massive biolab empire on Russia’s border, funded by American taxpayers.
Who authorized this? Why were these deadly pathogens there? And why did the media and officials deny it for years while calling it “Russian disinformation”?
This needs full Congressional investigation NOW. Heads must roll.
America deserves answers. The world deserves the truth.
145
1,975
2,724
27,230
THEE Gregory Stewart retweeted

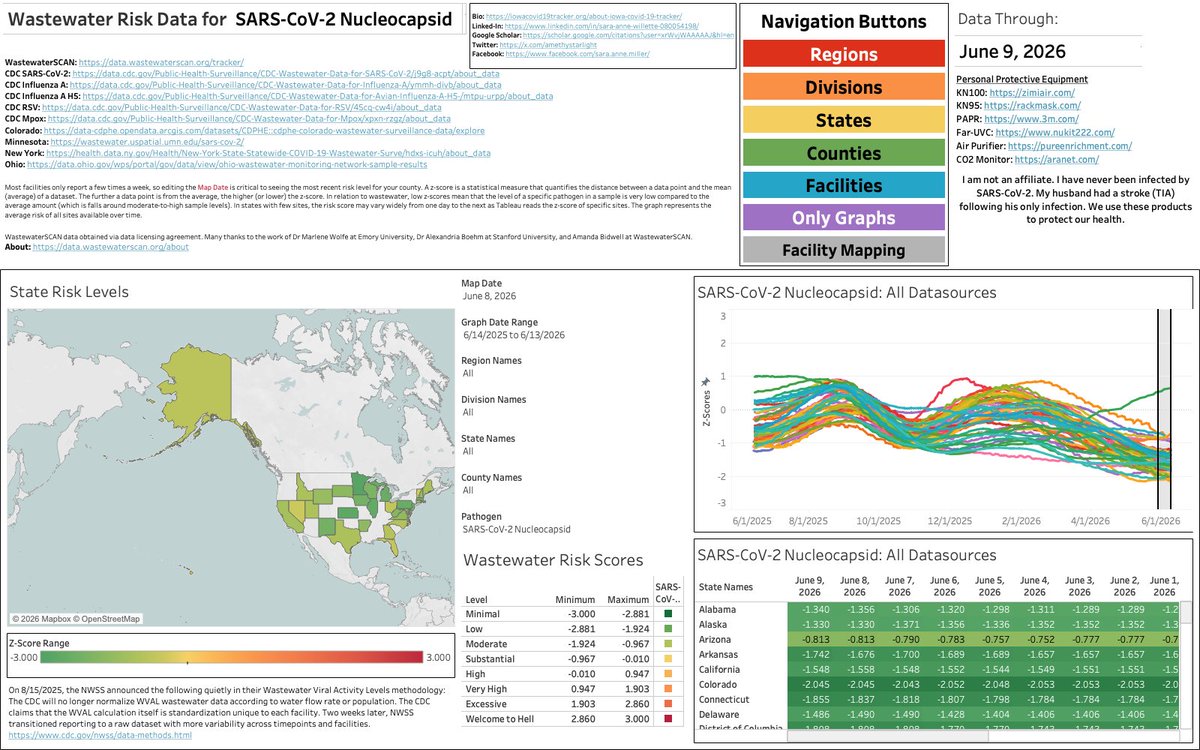
Throughout winter respiratory virus season and non-seasonal waves, I breakdown US wastewater trends across pathogens for each US region on Sunday nights. The next SARS-CoV-2 wave is beginning now because exponential increase appears slow at the start, nearly imperceptible. The CDC’s own epidemic growth likelihood model agrees with my analyses.
If you have important medical/dental things to handle and are in the Midwest or the Northeast, get it done now. The window for unmasked medical/dental care is closing for the South and the West.
The best things you can do for your health are:
Wear a fitted respirator in shared air spaces.
Clean indoor air with HEPA and far-UVC.
Rest at home when you are infectious and recovering.
Keep all your boosters up-to-date, including SARS-CoV-2 and MMR.
All visualizations are found at iowacovid19tracker.org. Data are subject to change as new data populate in state & federal repositories.

1
10
33
881
Kcplus3 🇺🇸 retweeted

Jun 12
autonomous robot driving through the field at night. no chemicals. no pesticides. just UV light killing pathogens and pests while everyone sleeps. this is @tricrobotics.
this is what chemical-free pest control looks like at scale.
1,437
10,726
63,015
5,750,416

OBAMA was involved in the CREATION of U.S. BIOLABS conducting gain-of-function research of deadly pathogens across the world, especially in Ukraine using U.S. taxpayer dollars, through agencies like USAID.
After the bioweapons convention ban ended in 2001 and after the anthrax attacks, Dr. Fauci essentially restarted the bioweapons arms race under the pretension of developing vaccines.
In 2014, Obama moved Fauci's gain-of-function research offshore after Obama was essentially begged to shut down operations from over 250 scientists because of the dangerous research Dr. Fauci was conducting.
President Trump was the one who shut down gain-of-function research in the U.S. and abroad.
DNI Tulsi Gabbard first exposed these labs in 2022 and now the proof of 120 biolabs in 30 countries, with a heavy focus on Ukraine, where the U.S. built and supported over 40 labs has been released and it all goes right back to the Obama administration and Dr. Fauci.
301
6,193
10,609
81,102
Tracy R retweeted

‼️What just happened?! - Nobody knows what the secret labs were for
That is the scariest part: the pathogens were real, the oversight failed, and no one can say how many more labs are out there.
Source: NewsForce
Host: @paddyney
10h

‼️What Just Happened?! ! The Secret Biolabs Found in America
America was watching the skies for Ebola. It should have been watching a warehouse in California.
@paddyney dives into the secret biolabs found in Reedley and Las Vegas, the CCP-linked fugitive behind them, and the biosecurity threat hiding in plain sight.
01:21 - A parking complaint exposed a secret lab
02:58 - The fugitive who did it twice
04:21 - Washington is panicking, both parties agree
05:26 - It wasn’t just two labs
06:49 - The invisible war
09:33 - Nobody can tell you why
1
9
16
5,141

The existence and purpose of many of these facilities had already been publicly documented for years. The U.S. Department of Defense stated that its Biological Threat Reduction Program in Ukraine was intended to strengthen disease surveillance, diagnostics, and biosafety, and to reduce threats posed by dangerous pathogens:
"Since 2005, BTRP has partnered with the Government of Ukraine to support peaceful and safe biological detection and diagnostic capabilities and to reduce the threats posed by pathogens."



1
6
R. Marshall Smith ☮️🎃 retweeted

"Many of these US government-funded biolabs were conducting research on dangerous and highly contagious pathogens, in some cases including gain-of-function research, with virtually no oversight or supervision," the Office of the DNI stated.
@cafreeland @MarkJCarney @AnitaAnandMP @nytimes @CdnPressNews #cdnpoli #Ukraine
Jun 12

🇺🇸🇺🇦 US Intel Declassifies Files on American Biolabs in Ukraine Researching Dangerous Pathogens
Outgoing Director of National Intelligence Tulsi Gabbard — set to leave her post at the end of the month — has published a tranche of declassified documents detailing the activities of US-funded biological laboratories in Ukraine.
"Many of these US government-funded biolabs were conducting research on dangerous and highly contagious pathogens, in some cases including gain-of-function research, with virtually no oversight or supervision," the Office of the DNI stated.
Gabbard added: "Despite the obvious potential for catastrophic global consequences that research on dangerous pathogens can have, policymakers and so-called medical officials — such as Dr. Fauci — along with figures in the Biden administration's national security team lied to the American people about the existence of US-funded and supported biological laboratories, and threatened those who tried to expose the truth. The Office of the Director of National Intelligence will continue to work closely with partners across government to determine where these labs are located, what pathogens they contain, and to put an end to dangerous gain-of-function research that threatens the health and wellbeing of the American people and people around the world."
The declassified documents reference several such facilities — including centers in Kharkov, Dnepropetrovsk, Lvov, Vinnitsa, and Chernigov — where research was conducted on anthrax, tularemia, swine flu, Marburg fever, Ebola, plague, and other pathogens.

1
3
109