
Rare earth elements power modern tech—but are hard to isolate. Joseph Cotruvo, Jr.’s (@PSUScience) discovery of lanmodulin, a protein that binds particular rare earth elements w/remarkable specificity, is changing that.
⏯️#ASBMB26 talk: youtu.be/8BWTQbIifmc #Lanthanides

ALT Joseph Cotruvo, Jr. (left), winner of the 2026 ASBMB Mildred Cohn Young Investigator Award, and J. Martin Bollinger Jr. (right) pose for photo holding the award at the 2026 ASBMB Annual Meeting.

ALT Joseph Cotruvo, Jr., winner of the 2026 ASBMB Mildred Cohn Young Investigator Award, stands at a podium delivering his talk at the 2026 ASBMB Annual Meeting.
4
8
582
Rare earth elements power modern tech—but are hard to isolate. Joseph Cotruvo, Jr.’s (@PSUScience) discovery of lanmodulin, a protein that binds particular rare earth elements w/remarkable specificity, is changing that.
⏯️Watch his #ASBMB26: youtu.be/uRvVuDv5d4I?si=bhFj….
#Lanthanides
2
7
737

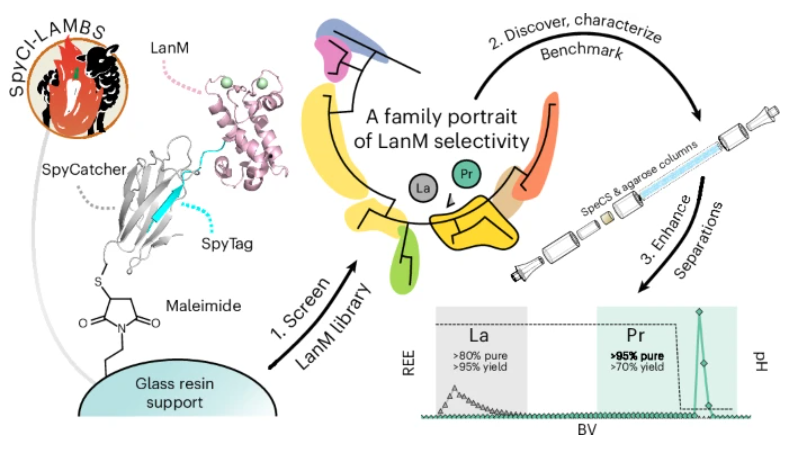
A new paper reports a high-throughput platform for quantifying the rare earth metal selectivity of lanmodulin proteins, enabling high-purity rare earth metal recovery and separations
nature.com/articles/s41589-0…
1
6
22
2,548

27 Oct 2025
⚛️ Is the secret to circular rare earth recovery hidden in molecular motion?
🔗Computationally derived structural insights into Rare Earth selectivity in lanmodulin and its variants. DOI: doi.org/10.1016/j.csbj.2025.…
📚CSBJ: csbj.org/
#RareEarthElements @CSB_Journal
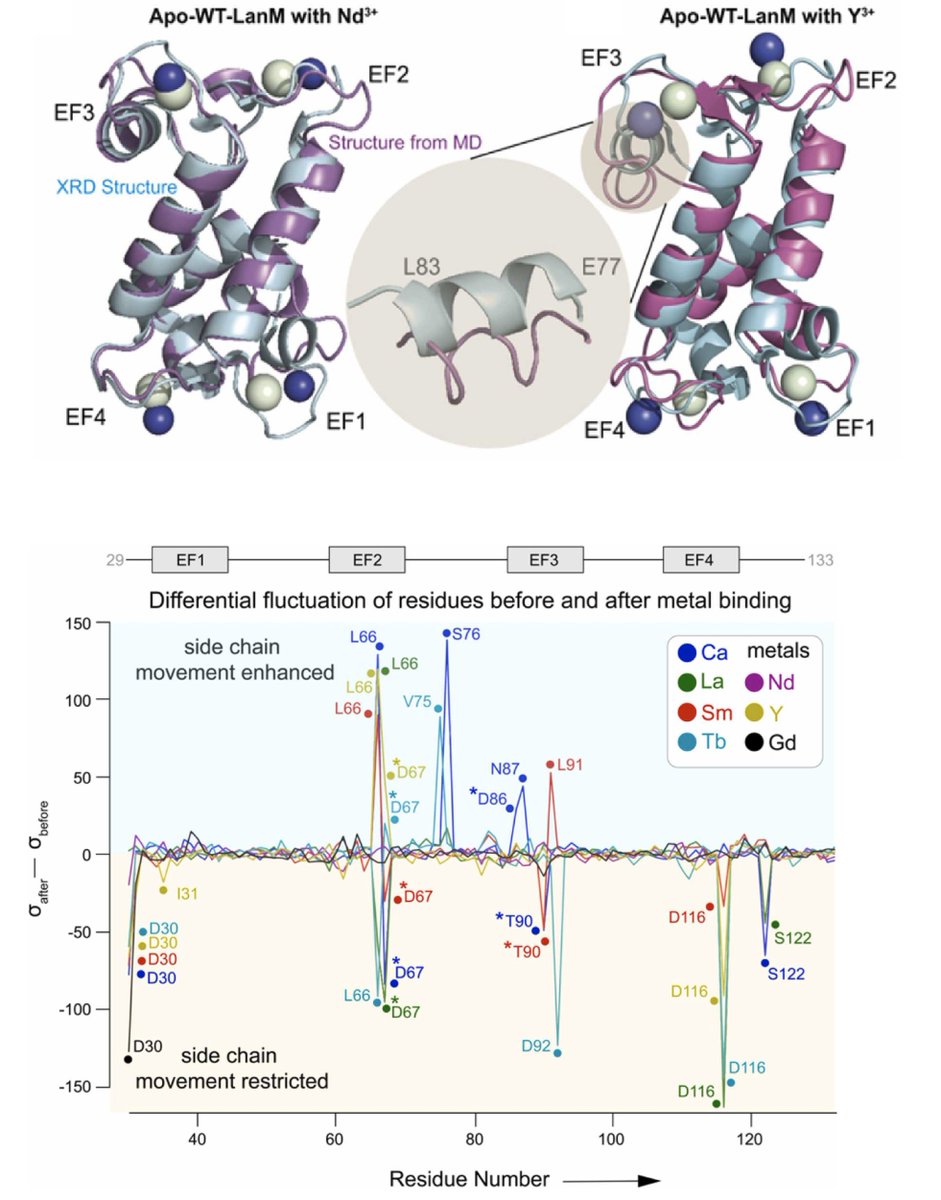
ALT Computationally derived structural insights into Rare Earth selectivity in lanmodulin and its variants. Computational and Structural Biotechnology Journal, DOI: https://doi.org/10.1016/j.csbj.2025.02.005
1
2
85

26 Jun 2025
check out @logandmorton Logan's latest paper in collaboration with @CotruvoLab published in @ACSBiomaterials on 'Mechanically and Chemically Robust Lanmodulin-Functionalized Silk Sponges for Rare Earth Element Sequestration' | @jugalsahoo @TuftsEngineer
pubs.acs.org/doi/10.1021/acs…
4
4
15
579

20 Jun 2025
The biomineralized protein formed by Lanmodulin -doped zinc imidazolate framework-8 for enhanced adsorption of rare earth elements sciencedirect.com/science/ar…
2
445

EF-Hand Battle Royale: Hetero-ion Complexation in Lanmodulin
By Dan M. Park et al. @Livermore_Lab @penn_state
Read the paper 👉 go.acs.org/c3r

2
468

15 Jun 2024
How does Gluconobacter oxydans dissolve minerals to get metals for energy technologies or CO2 sequestration? A new preprint by Sabrina Marecos in my lab tells us more about this.
biorxiv.org/content/10.1101/…
We've known for a long time that G. oxydans is really effective at dissolving minerals that metals needed for sustainable energy technologies by secreting a mineral-dissolving cocktail called a biolixiviant. A lot of the biolixiviant’s activity can be explained by its production of organic acids: Gluconobacter takes glucose and oxidizes it to gluconic acid (with a pH of ≈ 2.2).
In 2021, Alexa Schmitz (then a postdoc in my lab, now CEO of REEgen) screened our G. oxydans whole genome knockout collection with two acid production screens (using pH indicating dyes), and found a set of genes that controlled the production of acids. Knocking out the gene that codes for the membrane bound glucose dehydrogenase (mgdH), or code for enzymes that make its cofactor (the pqq and tld operons), reduce mineral-dissolution by between 65% (tldE) to well over 99.5% (mgdH).
doi.org/10.1038/s41467-021-2…
Alexa was able to use these results to engineer a strain of G. oxydans that has dramatically improved bioleaching of rare earths.
doi.org/10.1101/2023.02.09.5…
But, David Reed at Idaho National Lab showed that Gluconobacter’s biolixiviant is up to seven times more effective than gluconic acid alone the same pH (depending on how you measure it). This is suggested to us for a long time that there is more in the biolixiviant than just acid.
The fact that if you knock out acid production reduces mineral dissolution by > 99% tells us that whatever this thing is (or things are), that it doesn’t act additively with the acid, but synergistically. This makes sense when you look at the Pourbaix diagrams of minerals and metals (these are pH-redox phase diagrams that tell you under what conditions a metal is soluble): the phase transitions aren’t horizontal or vertical, they are diagonal. This suggests to us that if you change the redox potential (through a redox molecule like a flavin or a quinone, or by direct redox interaction through something like Cyt-2 or Mtc), or if you change the chemical potential (through a chelator like a siderophore or lanmodulin), then you can amplify the effectiveness of an acid dramatically.
But, if Gluconobacter’s biolixiviant does contain redox proteins or chelators (or both), what are they? Because this mechanism is not pH based it wouldn’t have shown up on our earlier pH indicator based acid-production screens. To solve this problem Sabrina turned to a direct mineral dissolution screen. This screen directly measures dissolution of a synthetic monazite mineral (the mineral that contains rare earths at the Mountain Pass mine, made by Megan Holycross in Earth and Atmospheric Sciences) with the lanthanide-chelating dye Arsenazo-III. This screen is sensitive to gene disruptions that change the pH and even those that don’t.
Sabrina found 89 genes important for bioleaching, 68 of which have not previously been associated with this mechanism. We conducted bench-scale experiments to validate the extraction efficiency of promising strains: 8 demonstrated significant increases in bioleaching by up to 111% (G. oxydans δGO_1598, a disruption of the gene encoding the orotate phosphoribosyltransferase enzyme PyrE), and one strain significantly reduced it by 97% (δGO_1096, a disruption of the gene encoding the GTP-binding protein TypA).
We think that the 8 genes whose disruption significantly increases bioleaching are responsible for regulating the non-acid mechanism of bioleaching, and that disrupting them takes the brakes off non-acid bioleaching (like knocking out phosphate signaling does for acid production). Furthermore, we think that that TypA might have something to do with secretion of the non-acid bioleaching molecules.
But, there’s no smoking gun or silver bullet for a chelator or redox molecule. Why haven’t we seen it? One answer is that we just missed it in the genetic screen. This is possible, but the knockout collection that Sabrina built is highly curated. Another answer is that there isn’t a single molecule, there are many that are responsible for bioleaching, and that that knocking out any single one doesn’t affect bioleaching to within the sensitivity of the Arsenazo-III assay. Time, hopefully, will tell which one it is.
But, the genes that we do have, have given us an additional roadmap for engineering G. oxydans to further improve its ability to dissolve minerals, and get at the metals we need for the sustainable energy infrastructure.
Supported by @ARPAE, The Link Energy Foundation, @CornellCALS, @AtkinsonCenter, @CornellEng.
6
470

26 Mar 2024
Proud to share our new article in @ACS_AMI! 🥳First authors Geeta Verma and Jacob Hostert worked with our collaborators @Duval_Lab, @rbgetman, and @GTChBE to characterize the rare earth element affinity of surface-bound peptides derived from Lanmodulin: pubs.acs.org/doi/10.1021/acs…
2
3
25
2,120

11 Mar 2024
Iowa State Engineer Ratul Chowdhury and his team are leading a green revolution in rare-earth metal extraction, utilizing data-driven design, Chowdhury aims to optimize the Lanmodulin protein for stable and efficient industrial extraction. Discover more: news.engineering.iastate.edu….

3
3
271

12 Dec 2023
This is #Christmas before Christmas: #actinides on the supplementary cover of ES&T.
#MyACSCover🎄☢️🎁
Collaboration @Livermore_Lab @penn_state.
@EnvSciTech @NNSAnews #chemtwitter @ACS4Authors @ACSPublications #nuclear #protein #Lanmodulin
pubs.acs.org/doi/10.1021/acs…

ALT Environ. Sci. Technol. 2023, 57, 49, 20830–20843. https://pubs.acs.org/doi/10.1021/acs.est.3c06033
1
3
323

Electron Paramagnetic Resonance, Electronic Ground State, and Electron Spin Relaxation of Seven Lanthanide Ions Bound to Lanmodulin and the Bioinspired Chelator, 3,4,3-LI(1,2-HOPO) (Eaton, Cotruvo, Jr., et al.) @CotruvoLab onlinelibrary.wiley.com/doi/…
12
1,773
13 Nov 2023
Start your week with a dose of #protein (plus #rocks and #radioactive elements...) 💪🏼☢️🪨!
New paper in @EnvSciTech from our team & @CotruvoLab.
@NNSANews @doescience #actinide #lanmodulin #rareearths #nuclear_waste
pubs.acs.org/doi/10.1021/acs…

How do radioactive materials behave in the environment? @Livermore_Lab and @penn_state scientists recently found that natural proteins, called lanmodulins, can make radioactive elements more prone to migrate from their initial location: livermorelab.info/49ukPeU
1
6
1,483

14 Aug 2023
At #ACS2023? Come learn about our collaborative @NSF ECO-CBET project with cool data from @Duval_Lab and @RennerResearch!
“Adsorbents functionalized with lanmodulin-derived peptides for REE separations”
TODAY! 2:30PM PT, Moscone south, Room 314

4
28
2,820

20 Jul 2023
🥳congrats to @s_gutenthaler who just won the poster prize 🏆 at #ICBIC in Adelaide @SBICofficial ! She presented her work on lanmodulin inspired peptides @ERC_Research 🤓

ALT Young scientists on a stage holding poster prize awards
3
3
35
2,561

30 Jun 2023
Researchers have taken a crucial step in their quest to purify lanthanides using a natural protein called lanmodulin (LanM), which could offer a more efficient and environmentally friendly approach to isolating the metals. fal.cn/3zxdZ
1
11
2,629

19 Jun 2023
Need RE Rare Earth element refining?
Bacteria can do it!
The Oak tree has more than acorns.
Quercus hansschlegelia enzyme Lanmodulin enzyme ( dimers ) can do the job
#biomade 1/
iflscience.com/bacteria-are-…
1
1
2
160
17 Jun 2023
Researchers have taken a crucial step in their quest to purify lanthanides using a natural protein called lanmodulin (LanM), which could offer a more efficient and environmentally friendly approach to isolating the metals. fal.cn/3zbwd
1
14
6,416
9 Jun 2023
Researchers have taken a crucial step in their quest to purify lanthanides using a natural protein called lanmodulin (LanM), which could offer a more efficient and environmentally friendly approach to isolating the metals. fal.cn/3yXvQ
3
17
5,311

8 Jun 2023
Nature "Enhanced rare-earth separation with a metal-sensitive lanmodulin dimer" doi.org/10.1038/s41586-023-0… と解説 nature.com/articles/d41586-0…
イオン半径など化学的性質が酷似しているランタノイド類を区別できる蛋白質 lanmodulin の構造。
1
12
1,974