
Disseminating knowledge about single-cell technology. Like/RT ≠ endorse
Joined April 2021
- Tweets 1,607
- Following 4,240
- Followers 6,364
- Likes 1,093
22 Photos and videos













The Single-Cell Ninjas retweeted

It’s well known that inflammation increases cancer risk, but how?
The answer: the epigenome "remembers" inflammation and primes stem cells for cancer.
Here is our paper: nature.com/articles/s41586-0…
And a special shoutout to the lead author @snaga13
A 🧵
15
130
568
42,627
The Single-Cell Ninjas retweeted
𝗦𝘁𝗼𝗽 𝗳𝗶𝘅𝗮𝘁𝗶𝗻𝗴 𝗼𝗻 𝗳𝗶𝘅𝗮𝘁𝗶𝗼𝗻.
There. I said it. The single-cell field has developed a collective obsession with PFA-based fixation as the answer to one of our most persistent operational headaches: how do you decouple sample collection from cell capture?
It's a real problem. Anyone who has run a multi-site clinical trial, managed a core facility with unpredictable sample flow, or tried to batch longitudinal timepoints knows the pain. Samples degrade. Biology drifts. The clock starts ticking the moment tissue leaves the body.
So, the field reached for the most familiar tool in the histology drawer — formaldehyde — and declared the problem solved. Except it isn't solved. It's traded.
Here's what nobody wants to say out loud: PFA fixation solves your logistics problem by destroying half your biology.
Let me be specific, because vague claims don't move science forward:
→ PFA cross-links cause RNA fragmentation. That's why probe-based hybridization had
to replace poly(dT) capture in fixed-cell workflows. That's not innovation — that's a workaround for chemistry-induced damage.
→ PFA alters protein conformation through methylene bridging. Surface epitopes get masked. If you're trying to do high-fidelity protein quantification alongside transcriptomics — the very thing that makes single-cell multiomics transformative — you've compromised your starting material before you even load the instrument.
→ PFA-protein-DNA cross-links obliterate native chromatin architecture. ATAC-seq on
PFA-fixed material? Incompatible. Multiomic ATAC gene expression? Incompatible.
That entire epigenomic axis — gone.
→ Full-length TCR/BCR profiling requires intact, full-length mRNA. PFA fragments it.
Paired chain immune repertoire recovery from fixed material? Not happening.
→ And because fragmented RNA needs probe-based rescue, you're locked into pre-designed panels for human and mouse only. Working with NHPs, rat models, organoids from non-standard species? Out of luck.
So what are we actually preserving here — the sample, or just the RNA-seq data?
This is the question the field needs to confront honestly. If your experiment only needs gene expression from fixed cells, PFA-based workflows are perfectly adequate. I won't pretend otherwise. They work. The data is good. But "adequate for scRNA-seq" is not the same as "ready for the future of single-cell
biology."
And the future...well, the future is unambiguously multiome. The papers being published today — the ones reshaping our understanding of tumour microenvironments, autoimmune mechanisms, vaccine responses, neurodegeneration — they don't profile one analyte. They integrate transcriptomes, surface proteomes, intracellular signalling states, chromatin accessibility, and immune repertoires. From the same cells. Simultaneously. You cannot do that from PFA-fixed material. This is not a limitation that better engineering will fix. It's a thermodynamic consequence of cross-linking chemistry. The
information is destroyed at the point of preservation.
The alternative already exists. Non-crosslinking cryopreservation approaches — and I'll disclose my interest here, I'm referring specifically to BD OMICS-Guard™ CRYO — take a fundamentally different path. No methylene bridges. No protein denaturation. No RNA fragmentation by design. Cells are preserved in a near-native state, cryostored for months, and when you're ready to process them, they behave like fresh cells. That means every downstream modality remains open: WTA, CITE-seq, ATAC-seq, TCR/BCR profiling, intracellular protein detection, flow cytometry. All from one preserved aliquot. And here's the part that should make every core facility director pay attention: because the cells come out in a near-native state, they're not locked to one platform. You preserve once, then choose your instrument — whatever fits your question. Platform-agnostic preservation. One reagent. Every modality. Every platform.
Thoughts? Pushback?
7
7
66
9,024
The Single-Cell Ninjas retweeted

We fix cells A LOT, since it is currently the main, proven path.
It does resolve headaches... but sure, it moves the pain to other body parts. I won't take the side of one product vs another without doing an actual test, but the points raised are real and innovations are welcome
1
2
5
1,058
The Single-Cell Ninjas retweeted
Spatial Stage - The Festival of Genomics Biodata & AI 2026
In a new collaboration between Festival Of Genomics (@FLGenomics) and @GESTALT_sp…we have crafted the Spatial Stage bringing to you the most exciting advances in spatial biology.
I will miss out Boston (yeah, I know…) but my out-of-the-chart-amazing co-founders @ioavlachos and @DrJasPlummer as well as the one and only friend and family member @katycns will be here to welcome you all.
festivalofgenomics.com/bosto…
1
9
10
1,255
The Single-Cell Ninjas retweeted

I'm leading a $6.5M trial at @PittTweet (w/ @WeillCornell & @UNC) testing a natural mitochondrial booster for anti-aging, funded by Otsuka. The other folks involved @mason_lab @RobSchwartzLab @NitDawg
Volunteers needed, please reach out if interested!!!
medschool.pitt.edu/news/univ…
2
4
12
1,607
The Single-Cell Ninjas retweeted
#AGTA2026 returns to Sydney (Roundhouse UNSW), Sept 1316!
Confirmed superstar speakers:
Braden Tierney
Jiwoon Park
Martin Smith
Lipin Loo
Tiffany Boughtwood
Justin Wong
Joseph Powell
Kim-Anh Lê Cao
Topics:
Population-scale genomics (clinic non-human)
Spatial and Sc omics, disease atlases
CRISPR, functional/regulatory genomics
AI for genomics and pathology
Emerging platforms and automation
#AGTA2026 #Genomics #SpatialOmics #CRISPR #SingleCellOmics #AiGenomics #FunctionalGenomics #ClinicalGenomics
@agtaGenomics
1
2
7
1,056
The Single-Cell Ninjas retweeted
Hack alert! 🚨
In vivo multiomic Perturb-seq with enhanced nuclear gRNA capture | bioRxiv biorxiv.org/content/10.64898…
4
11
1,565
The Single-Cell Ninjas retweeted

Mar 10
Spatial perturb-seq: single-cell functional genomics within intact tissue architecture
Scale up your in situ CRISPR screens
Out in @NatureComms
2
39
207
17,743

Hello single cellers and also spatial people!
The Single Cell Ninjas are back on the road and would like to offer 1:1 tele-consultations sessions to discuss experimental designs, technologies choices, troubleshooting and so on. All completely free of charge. Please DM us!
7
13
2,024
The Single-Cell Ninjas retweeted
Join me and these 4 super legends in the fields of single cell and spatial‑omics for a fast‑paced discussion on innovative benchmarking approaches that address delivering the deepest biological insights per dollar, tailored to your specific study design.


𝗕𝗲𝗻𝗰𝗵𝗺𝗮𝗿𝗸𝗶𝗻𝗴 𝗶𝘀 𝗕𝗿𝗼𝗸𝗲𝗻. 𝗟𝗲𝘁’𝘀 𝗙𝗶𝘅 𝗜𝘁.
Single-cell and spatial benchmarking studies dominate journals and conferences. Yet, after the leaderboards are published, you’re still left with the same high-stakes questions:
Do these metrics actually inform your next biological move, or just confirm what you already know? When the grant money is on the line, the question isn't just 'what works' - it’s 'what works within reach.' Which platform truly maximizes biological output for your specific budget?
Most benchmarking work misses the point. It ranks tools. It crowns winners. But it leaves you to carry the risk when it’s time to design your study and spend your grant.
𝗠𝗼𝘃𝗶𝗻𝗴 𝗕𝗲𝘆𝗼𝗻𝗱 𝘁𝗵𝗲 𝗟𝗲𝗮𝗱𝗲𝗿𝗯𝗼𝗮𝗿𝗱
At AGBT 2026, four of our beloved leaders in the field will challenge the status quo of benchmarking.
We aren't talking about which method "wins" or who runs the fastest pipeline. We are questioning the utility of the metrics themselves.
This discussion will dive into the hard truths of study design:
-𝗜𝗻𝘀𝗶𝗴𝗵𝘁 𝘃𝘀. 𝗠𝗲𝘁𝗿𝗶𝗰𝘀: are you optimizing for biological discovery or just "clean" data?
-𝗥𝗲𝗮𝗹𝗶𝘁𝘆 𝘃𝘀. 𝗜𝗱𝗲𝗮𝗹𝗶𝘀𝗺: are you planning studies real labs can afford, or chasing extreme depth that isn't reproducible?
-𝗚𝘂𝗶𝗱𝗮𝗻𝗰𝗲 𝘃𝘀. 𝗛𝘆𝗽𝗲: are benchmarks helping you design better experiments or simply feeding a hype cycle?
𝗝𝗼𝗶𝗻 𝘁𝗵𝗲 𝗖𝗼𝗻𝘃𝗲𝗿𝘀𝗮𝘁𝗶𝗼𝗻
If you generate or analyze single-cell or spatial data, this panel will speak directly to your results and your bottom line.
Details coming soon. Stay tuned!
#Benchmarking #SingleCell #MultiOmics #Bioinformatics #Genomic #SpatialBiology #AGBT2026
7
18
2,853
The Single-Cell Ninjas retweeted
𝗕𝗲𝗻𝗰𝗵𝗺𝗮𝗿𝗸𝗶𝗻𝗴 𝗶𝘀 𝗕𝗿𝗼𝗸𝗲𝗻. 𝗟𝗲𝘁’𝘀 𝗙𝗶𝘅 𝗜𝘁.
Single-cell and spatial benchmarking studies dominate journals and conferences. Yet, after the leaderboards are published, you’re still left with the same high-stakes questions:
Do these metrics actually inform your next biological move, or just confirm what you already know? When the grant money is on the line, the question isn't just 'what works' - it’s 'what works within reach.' Which platform truly maximizes biological output for your specific budget?
Most benchmarking work misses the point. It ranks tools. It crowns winners. But it leaves you to carry the risk when it’s time to design your study and spend your grant.
𝗠𝗼𝘃𝗶𝗻𝗴 𝗕𝗲𝘆𝗼𝗻𝗱 𝘁𝗵𝗲 𝗟𝗲𝗮𝗱𝗲𝗿𝗯𝗼𝗮𝗿𝗱
At AGBT 2026, four of our beloved leaders in the field will challenge the status quo of benchmarking.
We aren't talking about which method "wins" or who runs the fastest pipeline. We are questioning the utility of the metrics themselves.
This discussion will dive into the hard truths of study design:
-𝗜𝗻𝘀𝗶𝗴𝗵𝘁 𝘃𝘀. 𝗠𝗲𝘁𝗿𝗶𝗰𝘀: are you optimizing for biological discovery or just "clean" data?
-𝗥𝗲𝗮𝗹𝗶𝘁𝘆 𝘃𝘀. 𝗜𝗱𝗲𝗮𝗹𝗶𝘀𝗺: are you planning studies real labs can afford, or chasing extreme depth that isn't reproducible?
-𝗚𝘂𝗶𝗱𝗮𝗻𝗰𝗲 𝘃𝘀. 𝗛𝘆𝗽𝗲: are benchmarks helping you design better experiments or simply feeding a hype cycle?
𝗝𝗼𝗶𝗻 𝘁𝗵𝗲 𝗖𝗼𝗻𝘃𝗲𝗿𝘀𝗮𝘁𝗶𝗼𝗻
If you generate or analyze single-cell or spatial data, this panel will speak directly to your results and your bottom line.
Details coming soon. Stay tuned!
#Benchmarking #SingleCell #MultiOmics #Bioinformatics #Genomic #SpatialBiology #AGBT2026
4
39
6,009
The Single-Cell Ninjas retweeted

Jan 22
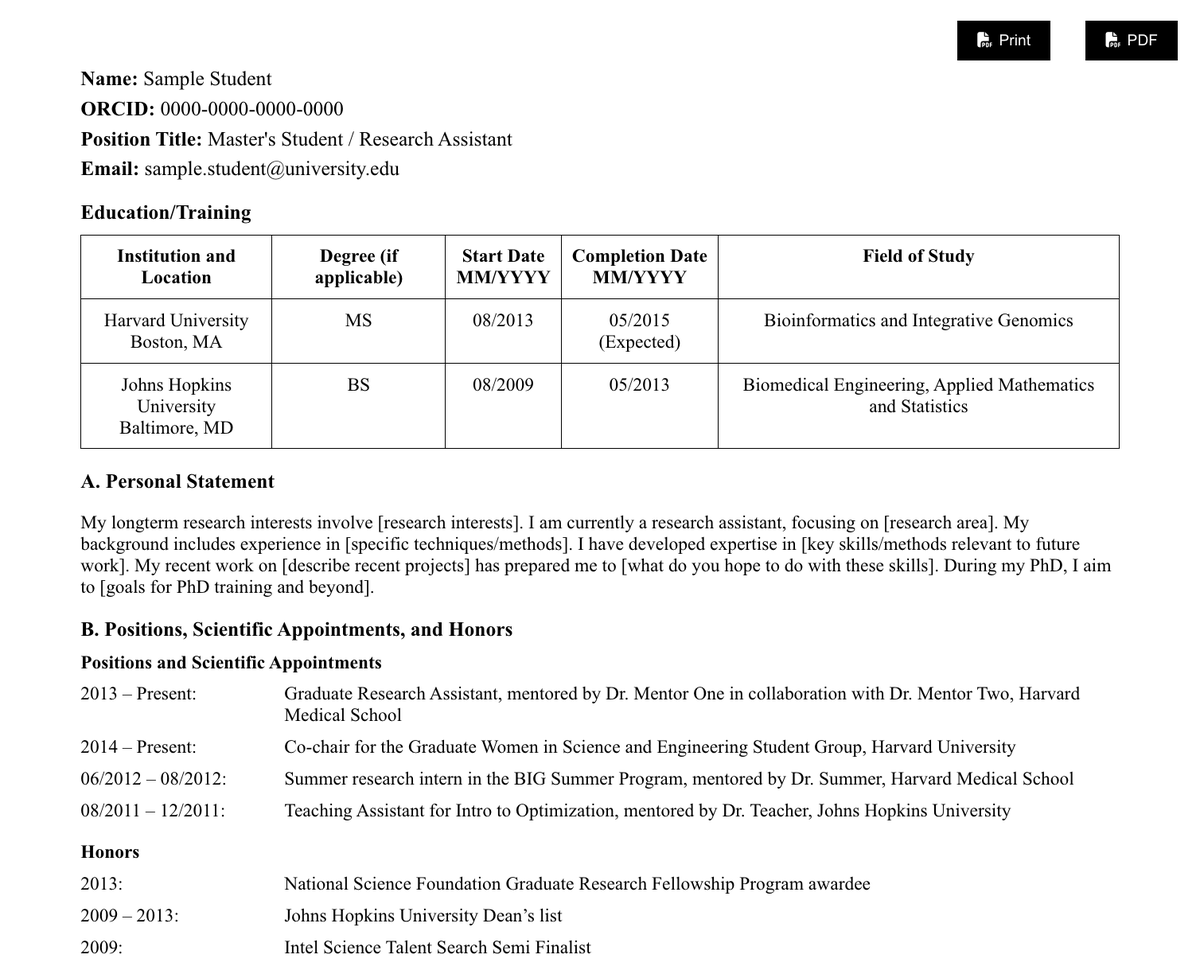
As I'm updating my NIH Biosketch for the N-th time this year, I made a Jekyll theme for students to familiarize themselves with this CV structure while building their online presence.
Demo: jefworks.github.io/online-bi…
Fork to modify: github.com/JEFworks/online-b…
#JustAcademicThings
2
18
125
15,933
The Single-Cell Ninjas retweeted
I just finished reading this beauty from the @KleinLabHMS 🫶🫶🫶
Multistep genomics on single cells and live cultures in subnanoliter capsules. Mazelis et al, recently published Science last December.
DOI: 10.1126/science.ady7209
I first saw Ignas start working on this almost five years ago when I was at HMS. It’s unreal, and makes me very happy, to see it go all the way to a Science paper. Legend!
In this work, Ignas and team introduce a new single-cell functional genomics platform that lets you:
-keep single cells alive
-culture them over time
-run multi-step biochemical workflows on them
-then read out genome-wide signals
The big breakthrough is the capsule design. It physically holds cells (and large biomolecules) in place, while still allowing media, enzymes, and reagents to diffuse in and out. Smart idea. Huge impact.
To make this work, they built “capsules with amphiphilic gel envelopes,” or CAGEs. These magic capsules:
-retain cells and large analytes (so you don’t lose the important stuff)
-stay permeable to media, enzymes, and reagents (so complex protocols still work)
-support high-throughput processing (so you can scale to tens of thousands of samples)
This matters because most functional genomic assays require multiple sequential steps. Those workflows fall apart when you try to scale them across large numbers of living single cells. CAGEs solve that, which allowed the team to:
-run high-throughput, multi-step workflows on living single cells (not limited to “one-pot” reactions)
-create a DNA library barcoding strategy for CAGE-derived material, so you can pool capsules without losing cell identity
-measure how gene expression programs persist through clonal expansion
-link lineage-like clonal growth with transcriptome readouts at real scale
Most single-cell sequencing methods are great at snapshots. But biology is dynamic. We care about things like:
-response to stimuli
-stability of cell states
-differentiation
-expansion of rare clones
This platform tackles one of the biggest bottlenecks in the field:
Scaling live-cell, multi-step functional measurements to high throughput, while still getting genome-wide readouts!!!
Big congratulations to the entire team!
I remember seeing the very first tests in this magical work…
Amazing science from @KleinLabHMS
Multistep genomics on single cells and live cultures in subnanoliter capsules | Science science.org/doi/10.1126/scie…
3
14
4,127
The Single-Cell Ninjas retweeted
Amazing pub from an even more amazing team! Congratulations to everyone!

TY @Aiims1742!
Thrilled to share our new @AGA_CMGH paper!
Using FixNCut scRNA-seq, we uncover novel acinar metaplastic cell states and precancer programs in pancreas disease.
sciencedirect.com/science/ar…
Led by the amazing @Aney_Katie and Woo-Jeong 👏🧵👇
1
3
1,276
The Single-Cell Ninjas retweeted

TY @Aiims1742!
Thrilled to share our new @AGA_CMGH paper!
Using FixNCut scRNA-seq, we uncover novel acinar metaplastic cell states and precancer programs in pancreas disease.
sciencedirect.com/science/ar…
Led by the amazing @Aney_Katie and Woo-Jeong 👏🧵👇
27 Dec 2025

New paper in @AGA_CMGH from @SaharNissim
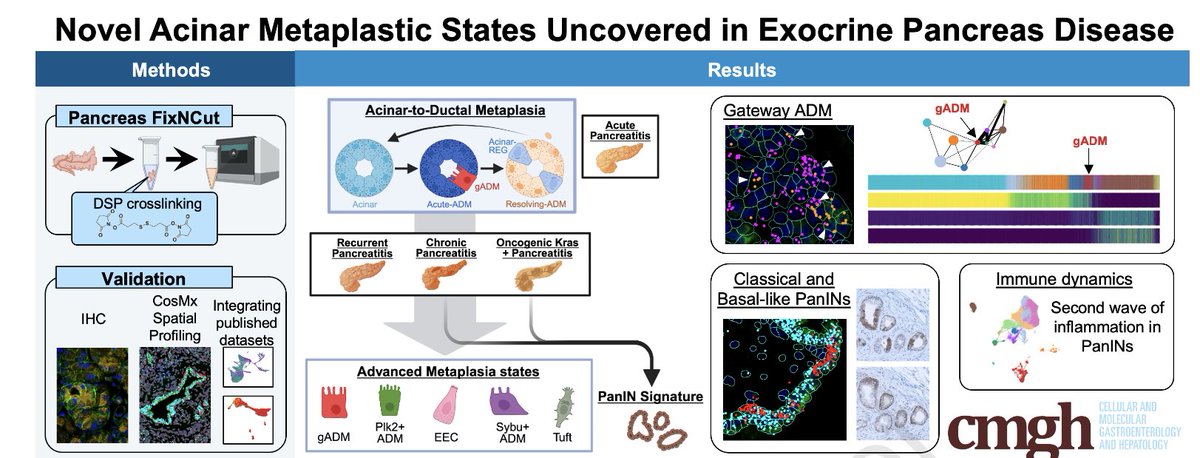
Novel Acinar Metaplastic States Uncovered in Exocrine Pancreas Disease
sciencedirect.com/science/ar…
Using FixNCut (which preserves RNA integrity in the pancreas), this study explores acinar plasticity & lineage state heterogeneity in PanINs.
1
2
6
1,967
The Single-Cell Ninjas retweeted

5 Dec 2025
Early Spatial Adopters Develop Standards, Resources for Quality Control and Data Benchmarking | @GenomeWeb genomeweb.com/sequencing/ear…
7
9
1,795
The Single-Cell Ninjas retweeted

3 Dec 2025
Single Cell Imaging is the new Single Cell Sequencing. The very first STAMP at CCI following El Maestro. STAMP in the quest to Zero Childhood Cancer. @LGMartelotto @DrJasPlummer @CytoNology @KidsCancerInst

4
8
1,074
The Single-Cell Ninjas retweeted

3 Dec 2025
Goliath effort to help with standards in the field of spatial trancriptomics … can only be more amazing when done with @LGMartelotto @mason_lab 🚀
Paper: nature.com/articles/s41587-0…
Touchstone portal: spatialtouchstone.org
Spatial touchstone protocols: protocols.io/view/spatial-to…
3 Dec 2025

A collaborative project led by St. Jude has established a comprehensive framework for standardizing spatial transcriptomics. The Spatial Touchstone project integrates publicly available imaging data with newly curated datasets across six tissue types. ow.ly/Ghrg50XBkr6
1
6
13
2,411
The Single-Cell Ninjas retweeted
3 Dec 2025
What started with three amigos’ shared frustration with weak quality metrics and SOPs in spatial omics grew into a framework built to help the community raise standards.
From frustration to focused progress.
@mason_lab @DrJasPlummer
stjude.org/media-resources/n…
10
20
3,959
The Single-Cell Ninjas retweeted
3 Dec 2025
Very happy to finally present our work!
“𝘚𝘵𝘢𝘯𝘥𝘢𝘳𝘥𝘪𝘻𝘦𝘥 𝘮𝘦𝘵𝘳𝘪𝘤𝘴 𝘧𝘰𝘳 𝘢𝘴𝘴𝘦𝘴𝘴𝘮𝘦𝘯𝘵 𝘢𝘯𝘥 𝘳𝘦𝘱𝘳𝘰𝘥𝘶𝘤𝘪𝘣𝘪𝘭𝘪𝘵𝘺 𝘰𝘧 𝘪𝘮𝘢𝘨𝘪𝘯𝘨-𝘣𝘢𝘴𝘦𝘥 𝘴𝘱𝘢𝘵𝘪𝘢𝘭 𝘵𝘳𝘢𝘯𝘴𝘤𝘳𝘪𝘱𝘵𝘰𝘮𝘪𝘤𝘴 𝘥𝘢𝘵𝘢𝘴𝘦𝘵𝘴”
Spatial transcriptomics promises deep insight into tissue architecture - but inconsistent data quality across labs and platforms has been a major barrier. A new study from a global consortium addresses this by releasing the Spatial Touchstone (ST): a harmonized, multi-site, multi-platform dataset using six tissue types and two widely imaging used technologies.
We introduce SpatialQM, an open-source quality-control software, and a public repository, Spatial Touchstone Portal (STP), that hosts ~33M cells and ~7B transcripts.
With standardized metrics for sensitivity, reproducibility, signal-to-noise, false discovery rates, and cell-type annotation, this framework gives labs a way to benchmark and compare spatial transcriptomics data reliably.
This effort sets a foundation for more robust, reproducible spatial-omics research — and helps unlock cross-study comparability across institutions and platforms.
I’m incredibly thankful to my amazing collaborators across the globe and an special shout out to my dear friends @DrJasPlummer and @mason_lab and their super amazing teams for literally working they brains out to make this happen.
Felipe, Jiwoon , David, Luke, Maycon, Yutian, Arjumand, Hannah, Kellie, Alex, Lisa, Alicia, Roberto and many more!
@WeillCornell
@StJudeResearch
nature.com/articles/s41587-0…
3
30
104
15,394