
The Benito-Gutierrez Lab uses amphioxus as a model system to understand how vertebrates evolved and conquered water, land and air
Joined March 2020
- Tweets 222
- Following 126
- Followers 237
- Likes 106
16 Photos and videos










AmphiSpace retweeted

23 Jun 2024
Arrived in Finland and super excited for the start of #EED2024 and the amphioxus satellite meeting!
In the meantime, recovering from jetlag in a very sunny Helsinki! 😎
20 Jun 2024

No big deal, just the view of Helsinki's senate square from #EED2024 as the organizers did our final walk-through today 😎 We are ready for the satellite meetings on Monday!

1
20
1,408

2 Feb 2024
Excited to see last Giacomo's @gattoni_giacomo work in the bioRxiv! Fun collaboration with Silvia Mercurio at Roberta Pennati's lab. Thank you all for your hard work, and giving the entire world another beautiful model system to work with 👏😀
1 Feb 2024

Very excited to share this new paper, fruit of a great collab with Silvia Mercurio, out in preprint! 🎉
We set up new methods for in vitro culture and gene/protein detection in the feather star A. mediterranea and used them to investigate its development. ⭐️ Tweetorial below!
2
418
AmphiSpace retweeted
1 Feb 2024
This feather star has it all: beautifully colored body, 10 arms, great at regeneration. Plus, it’s a #crinoid, the sister group to other living #echinoderms (starfish, sea urchin..), making it an ideal comparative system to understand how the puzzling echinoderm body plan evolved

2
1
5
325
AmphiSpace retweeted
1 Feb 2024
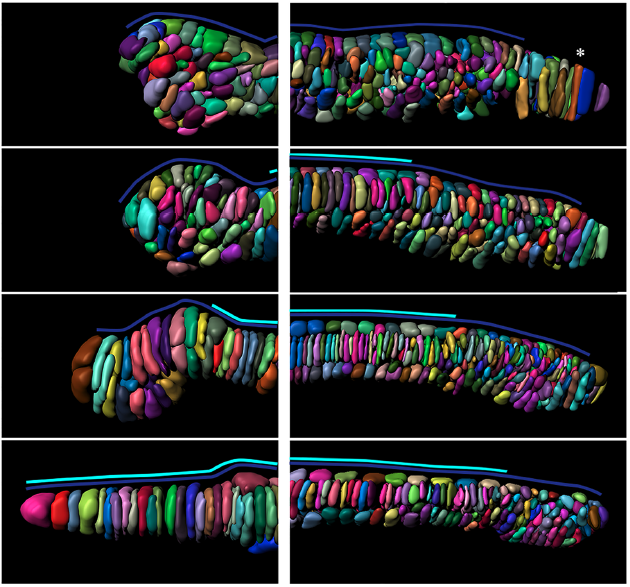
We then optimized whole-mount immunofluorescence, which allowed us to study how the different tissues of the crinoid #larva develop. Here for example we labelled the skeleton (yellow) and cilia (green, note the five ciliary bands and apical tuft!) of the doliolaria larva
1
1
2
249
AmphiSpace retweeted
1 Feb 2024
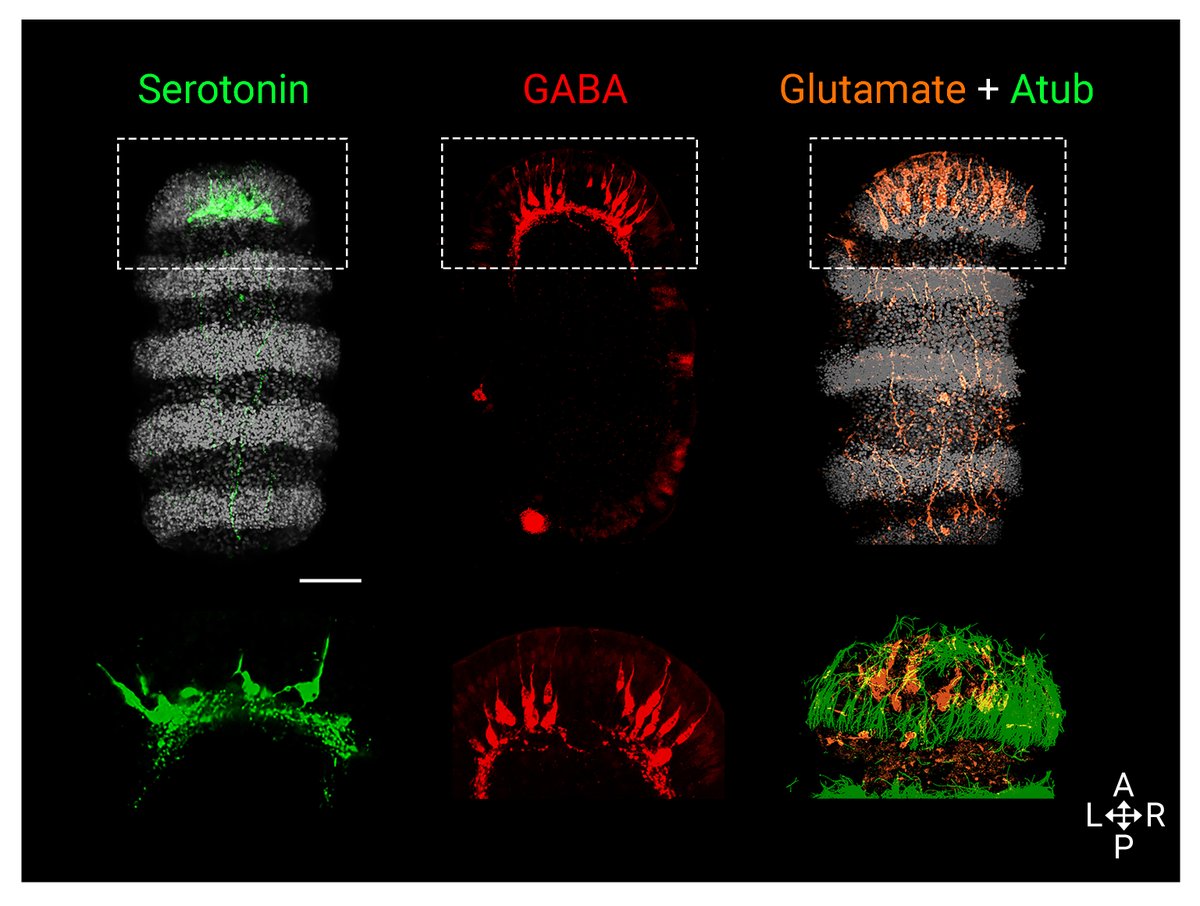
Next, we focused on our favorite, the nervous system, and looked at the distribution of serotonin, GABA and glutamate 🧠. We characterized the complexity of the larval apical organ (AO), and identified glutamatergic cells in the neural plexus
1
1
217
AmphiSpace retweeted
1 Feb 2024
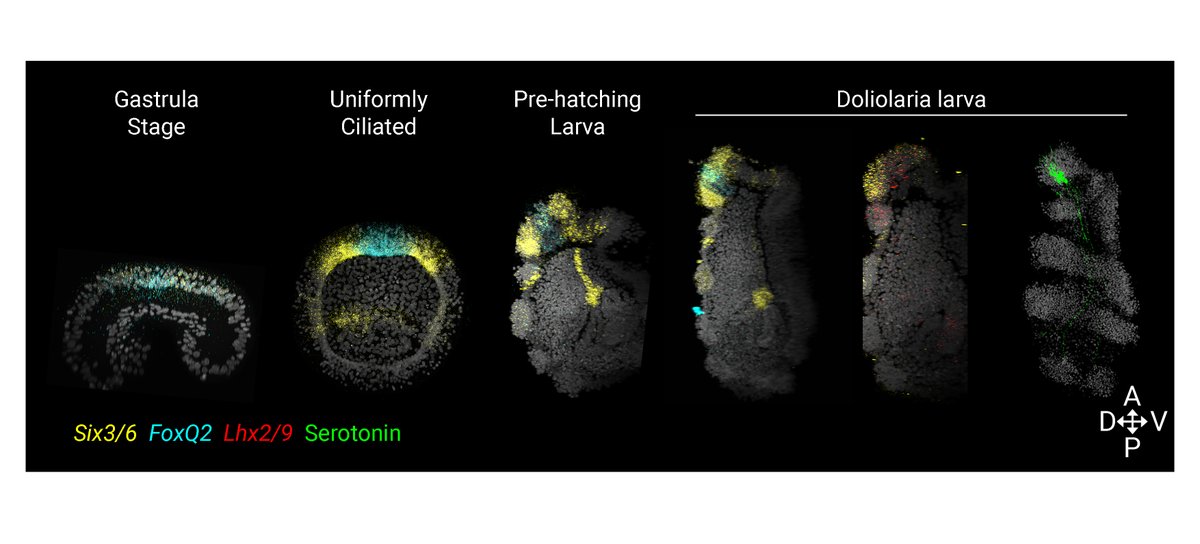
We then optimized in situ HCR (@HCRimaging) to test the molecular patterning of the crinoid AO. 🧬 We found that serotonergic neurons form in a domain co-expressing Six3/6 and FoxQ2 (from early development) and Lhx2/9 as in other echinoderms, supporting AO conservation
1
1
3
228
AmphiSpace retweeted
1 Feb 2024
At metamorphosis, larvae settle from the anterior pole, neurons degenerate and a new nervous system develops. We showed that cells in the oral/vestibular ectoderm, where the ectoneural system will form, start to express anterior markers in the post-metamorphic cystidean stage!
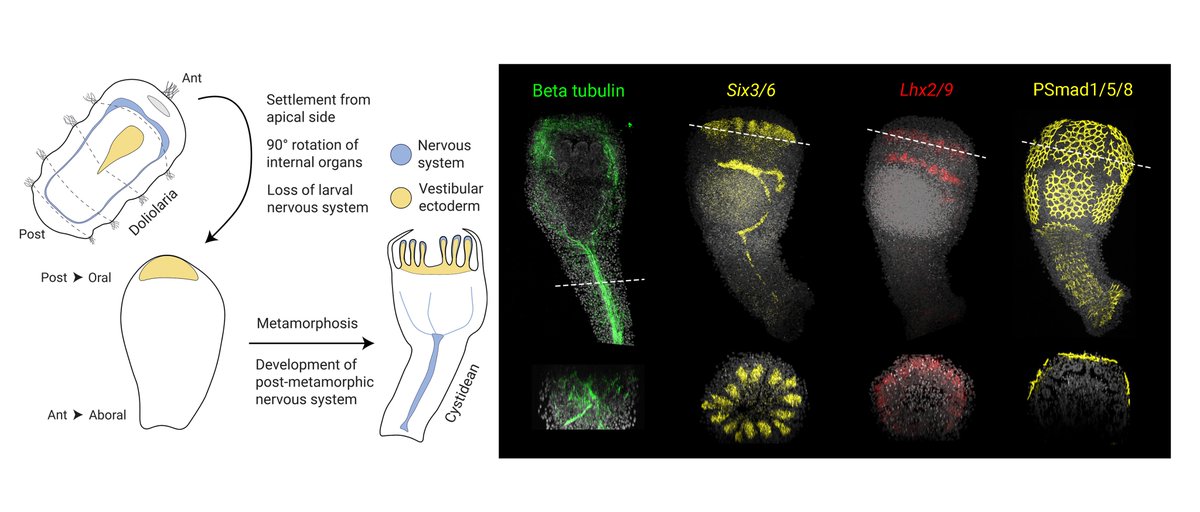
1
1
1
243
AmphiSpace retweeted
1 Feb 2024
This work has been an amazing collaboration between my PhD @CamZoology and Master’s @LaStatale labs, and I want to thank Silvia, both supervisors @eBGLab and Roberta Pennati, @MauriceElphick for all his help and support, all the authors, and all members of both labs! 😄
1
2
3
453
AmphiSpace retweeted

22 Jun 2023
🎁Interested in learning more about the study? Read also the Research Briefing:
rdcu.be/dfalY
nature.com/articles/s41556-0…
1
4
2,980
AmphiSpace retweeted
22 Jun 2023
☕️NEW: Ton, Keitley et al. provide a rabbit development atlas and propose that combining rabbit and mouse atlases can help dissect early primate development.
@mailinh_ton @DanKeitley @MarioniLab @BertieGottgens @eBGLab
rdcu.be/dfalo
nature.com/articles/s41556-0…
2
5
12
8,399
AmphiSpace retweeted

27 Apr 2023
Proudly presenting our new bioRxiv:
The cell type atlas of the regenerating and asexually reproducing annelid Pristina leidyi
We show that a piwi cell population with a pluripotent stem cell signature underlies adult differentiation
biorxiv.org/content/10.1101/…
A tweetorial 🧵👇
9
82
215
63,035
AmphiSpace retweeted
15 Mar 2023
I want to thank my wonderful (and patient) labmate @DanKeitley, all the @eBGLab, @CamZoology, @CRUK_CI and the Cambridge #EvoDevo community for their help and support!
If you have any questions/comments, please let us know! 😄
2
6
576
AmphiSpace retweeted
15 Mar 2023
Our work shows that an aGRN patterning the anterior neurectoderm is ancestral to bilaterians. In the chordate lineage the aGRN was incorporated into neurulation to specify and position the forebrain anteriorly, linking the evolution of the AO to that of the chordate brain.
10/10
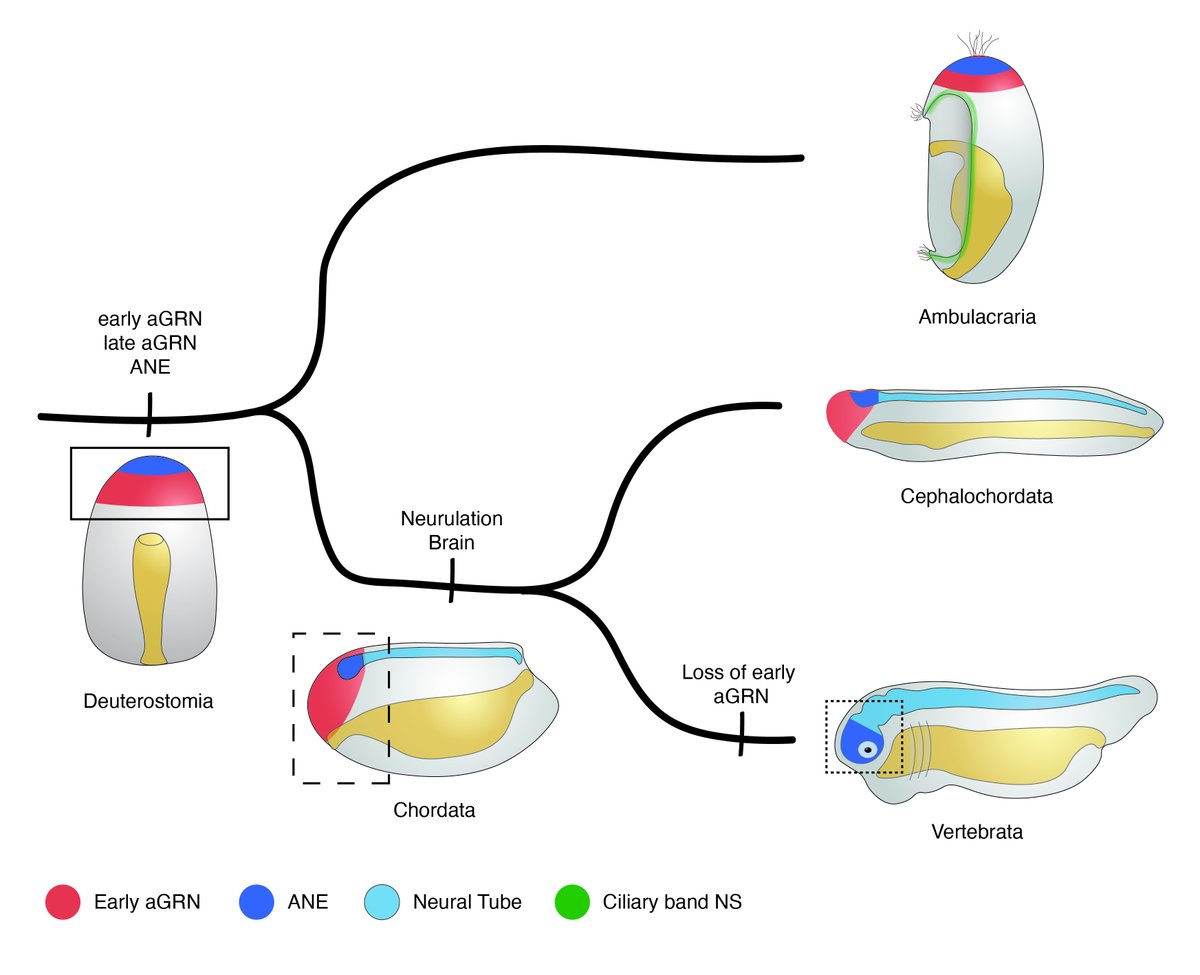
1
2
2
705
AmphiSpace retweeted
15 Mar 2023
But what exactly is this anterior identity? Here we describe a hypothalamic-like region in the amphioxus larval brain expressing Otp, FoxD, Bsx, Hmx, miR-7 and calcitonin-type neuropeptides. It forms in the area where aGRN is active and is lost following Wnt overactivation.
9/10
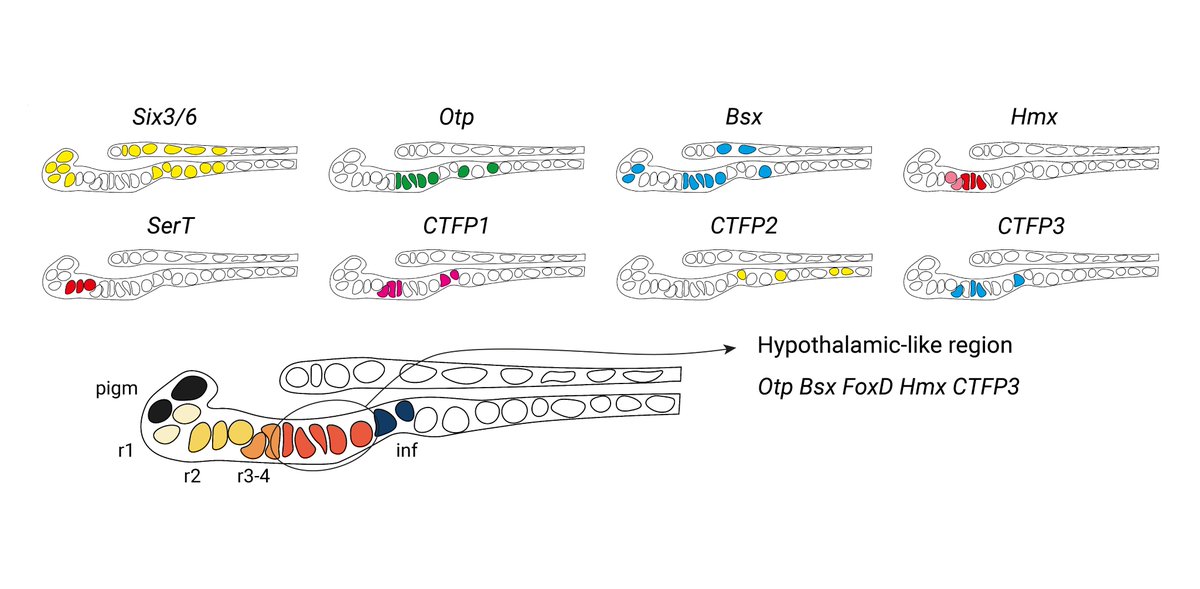
1
2
2
778
AmphiSpace retweeted
15 Mar 2023
As late Wnt overactivation does not lead to loss of neural cells, we can examine what happens to the brain when the aGRN is removed! We find that the aGRN is necessary to confer anterior identity to the neural plate, as the brain is posteriorized when the aGRN is repressed.
8/10
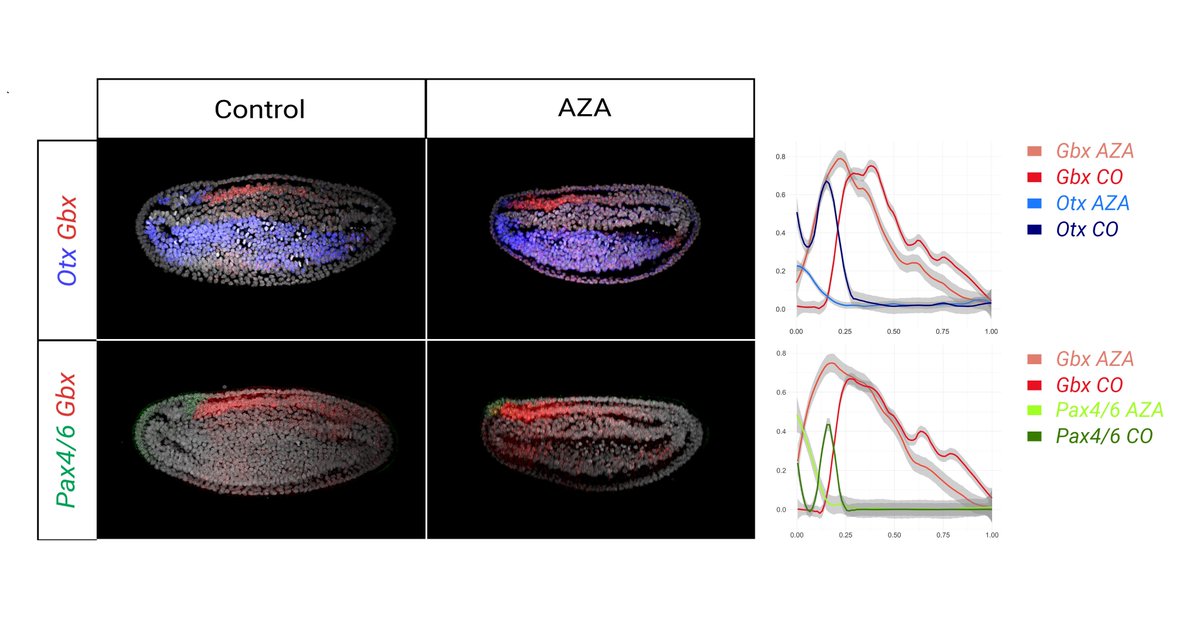
1
2
1
429
AmphiSpace retweeted
15 Mar 2023
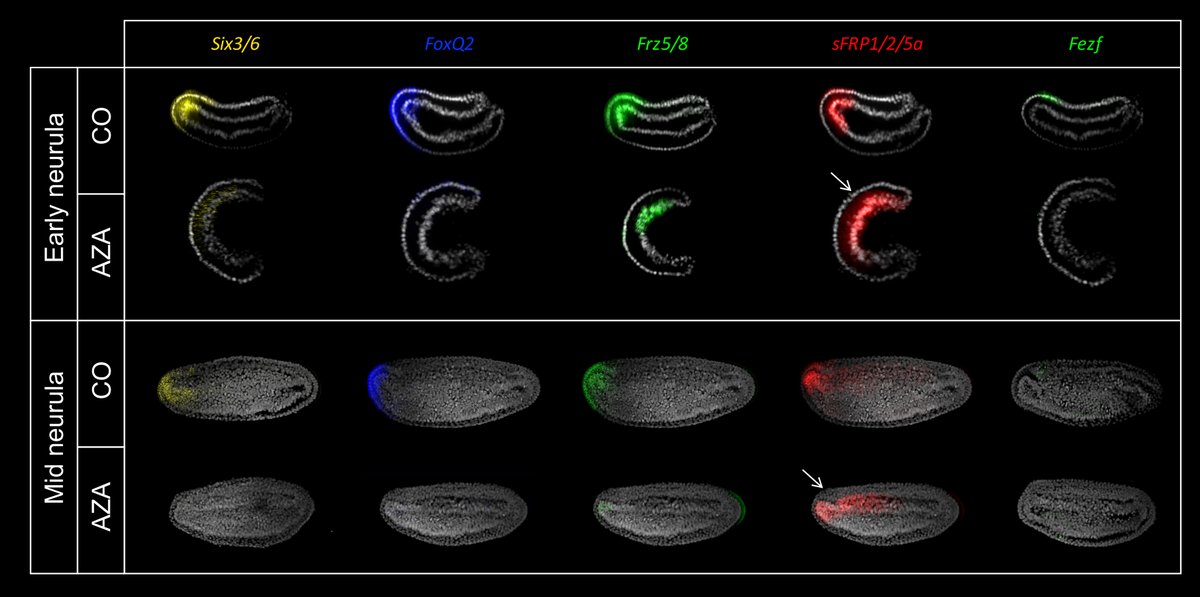
These results indicate that the aGRN is conserved in chordates! 🤩 To test whether the amphioxus aGRN is also controlled by Wnt, we overactivated Wnt signalling with Azakenpaullone and saw a downregulation of all aGRN markers in the ectoderm.
7/10
1
2
1
376
AmphiSpace retweeted
15 Mar 2023
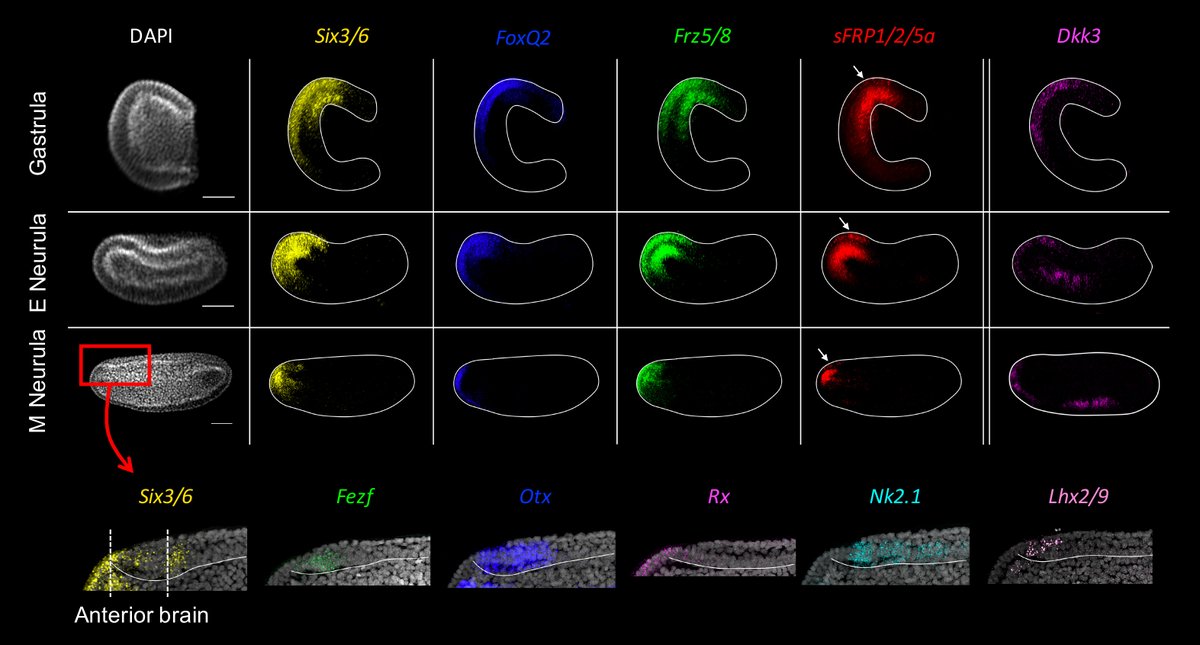
Investigation of aGRN genes in amphioxus revealed that early markers are co-expressed on the animal side and restrict anteriorly, while late markers appear in the anterior brain during neurulation, with a similar dynamic to apical organ development.
6/10
1
2
1
368
AmphiSpace retweeted
15 Mar 2023
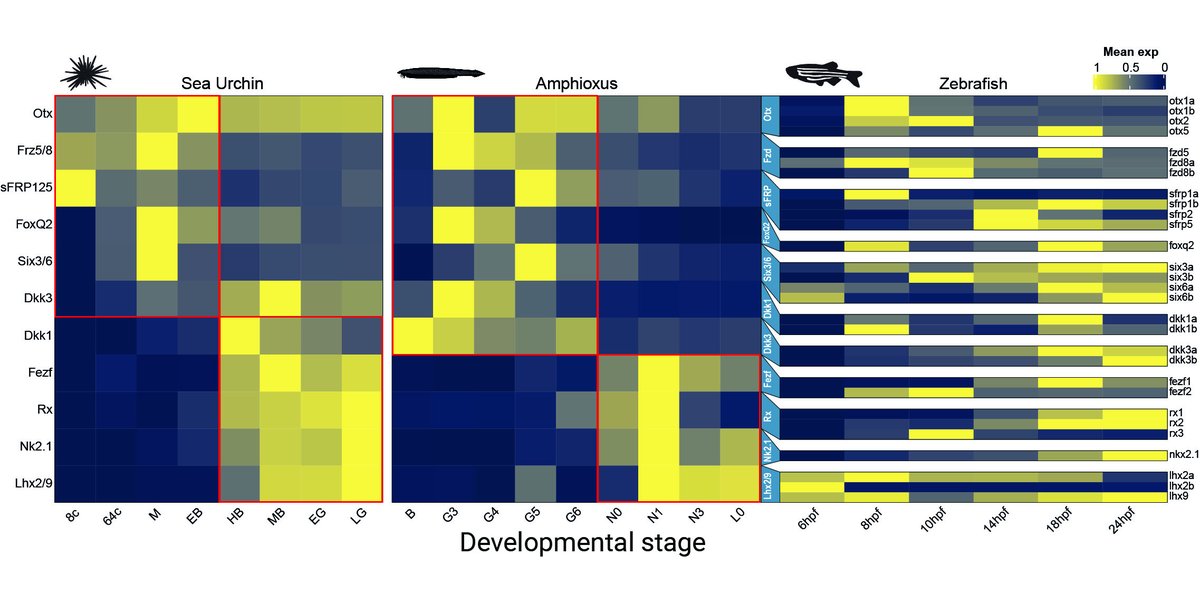
By comparing published scRNAseq datasets we find a biphasic activation of aGRN genes in sea urchin and amphioxus but in not zebrafish, where FoxQ2 is missing and the other markers are expressed later in the hypothalamus. This supports aGRN conservation in deuterostomes.
5/10
1
2
2
383
AmphiSpace retweeted
15 Mar 2023
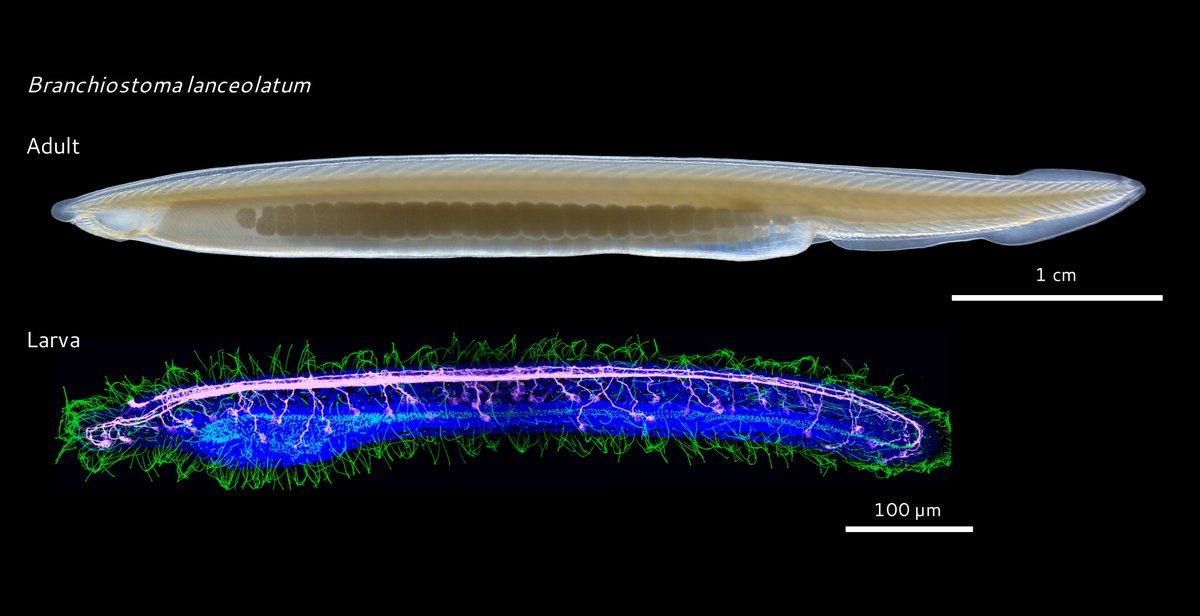
Is there a link between apical organs and chordate brains? To answer this question, we investigated the evolution of the aGRN in deuterostomes and traced its conservation in chordates using our favourite #amphioxus, a basally-branching chordate.
4/10
1
2
8
495
AmphiSpace retweeted
15 Mar 2023
Apical organ development is controlled by an anterior network (aGRN) composed of early repressive interactions between vegetal Wnt and animal Six3/6 and FoxQ2, which drive downstream apical organ genes. Many of the same genes are expressed in the vertebrate brain too!
3/10
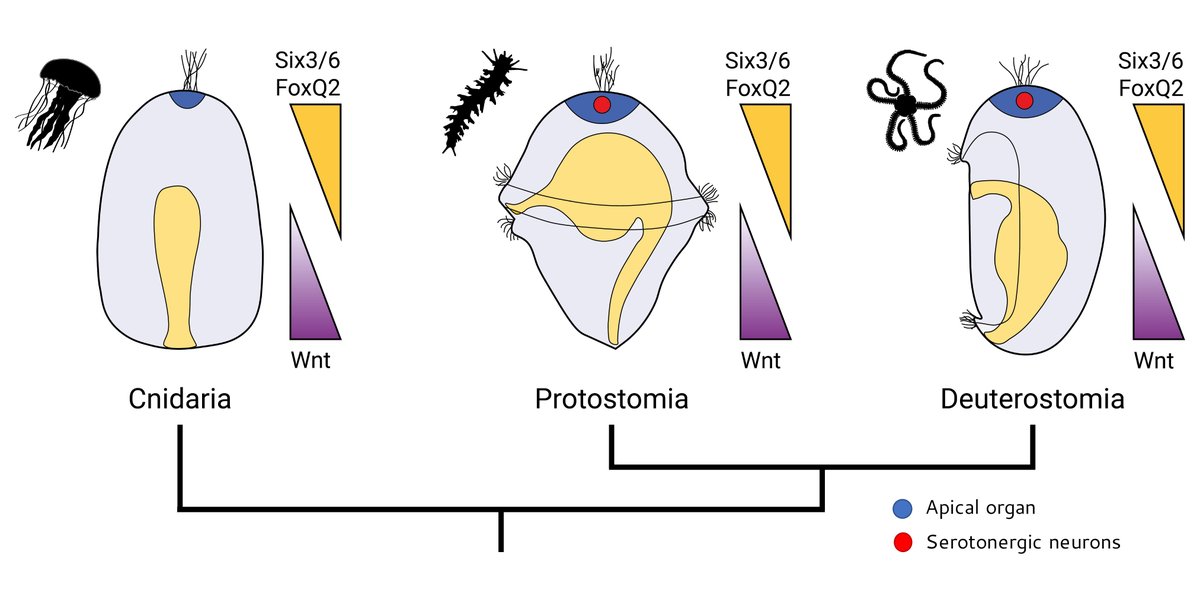
1
2
7
421