X retweeted

Jun 11
Have you seen our #ImageBank? 🤔📸
Our Image Bank forms a gallery of free-to-use images to ensure accurate, non-biased, and respectful images are widely available worldwide.
➡️ Find it here: worldobesity.org/resources/i…
#EndWeightStigma #PeopleFirst




1
1
96

Jun 11
imagebank別売り、ケーブルなし
imagebankと合わせて400GBPかぁ、ケーブルは専用よね、hasselbladrepairにあったっけ?
(そもそもimagebank使いたいかと言うとですね、という話に⋯)
32

MAEA potwierdza uszkodzenie elektrowni jądrowej w Zaporożu
Dziś rano ekipa Międzynarodowej Agencji Energii Atomowej (MAEA) potwierdziła uszkodzenia kontrolowanej przez Rosję elektrowni jądrowej w Zaporożu. MAEA zaobserwowała uszkodzenia metalowego włazu, a także kawałki złomu i spalone resztki światłowodu na ziemi. Zniszczenia, jak twierdzą, wskazują na atak drona.
Poziom promieniowania radioaktywnego nadal utrzymuje się w normie.
O atak Rosjanie oskarżyli stronę ukraińską. Wczoraj ukraińskie wojsko zaprzeczyło, jakoby był to ukraiński dron.
Elektrownia jądrowa w Zaporożu została przejęta przez Rosję w marcu 2022 roku
Fot. IAEA Imagebank

1
12
560
May 20
Have you seen our #ImageBank? 🤔📸
Our Image Bank forms a gallery of free-to-use images to ensure accurate, non-biased, and respectful images are widely available worldwide.
➡️ Find it here: worldobesity.org/resources/i…
#EndWeightStigma #PeopleFirst




2
172

Más de 1.000 imágenes de hemopatías accesibles desde su dispositivo móvil. #AtlasGECH #CitologíaHematólogica #Hematología #ImágenesMédicas #DiagnósticoMédico #GECH #FormaciónMédica #Hematopathology #ImageBank


18
41
2,437
Apr 23
Have you seen our #ImageBank? 🤔📸
Our Image Bank forms a gallery of free-to-use images to ensure accurate, non-biased, and respectful images are widely available worldwide.
➡️ Find it here: worldobesity.org/resources/i…
#EndWeightStigma #PeopleFirst




3
225
Mar 16
Have you seen our #ImageBank? 🤔📸
Our Image Bank forms a gallery of free-to-use images to ensure accurate, non-biased, and respectful images are widely available worldwide.
➡️ Find it here: worldobesity.org/resources/i…
#EndWeightStigma #PeopleFirst




3
211

Without justice systems that work for women, rights remain promises that never arrive.
This #IWD, we push for Rights. Justice. Action. For ALL women and girls. No stepping back.
#SwedenInNigeria #InternationalWomensDay
📸: Cecilia Larsson Lantz / ImageBank Sweden

1
3
445

King Letsie III and Queen 'Masenate Mohato Seeiso of of Lesotho celebrate their 26th wedding anniversary today!
Photo: IAEA Imagebank - CC BY-SA 2.0

3
8
247
10 Dec 2025
Have you seen our #ImageBank? 🤔📸
Our Image Bank forms a gallery of free-to-use images to ensure accurate, non-biased, and respectful images are widely available worldwide.
➡️ Find it here: worldobesity.org/resources/i…
#EndWeightStigma #PeopleFirst




2
218

How Weak Magnetic Fields Could Nudge Red Blood Cells into Clumping
When you look at blood under a microscope, healthy red blood cells (RBCs) normally glide past one another as separate, disk‑shaped cells. In many disease states, though, they start stacking up like coins. This reversible clumping is called rouleaux formation, and it thickens the blood, slows micro‑circulation, and changes how tissues are perfused.
Most explanations focus on plasma proteins such as fibrinogen and immunoglobulins. Those proteins screen the normal negative surface charge on RBCs, reducing the electrostatic repulsion that usually keeps cells apart. imagebank.
Here we explore a complementary idea: could weak magnetic fields influence the way red blood cells behave by subtly changing the spin states of electrons in their heme groups, and thereby nudging their surface charge and zeta potential?
This is not a claim that every phone call clots your blood. It is a mechanistic “what if?” grounded in what we already know about:
RBC electrostatics and zeta potential
The staggering number of heme groups in each cell
Spin chemistry and radical‑pair mechanisms in weak magnetic fields
1. Red Blood Cells, Zeta Potential, and Why They Don’t Normally Stick
Zeta potential: a small voltage with big consequences
Every RBC carries a slightly negative electrical potential at its surface, largely due to sialic acid residues in membrane glycoproteins. This surface charge creates an electrical double layer in the surrounding plasma and generates a zeta potential that keeps cells from sticking together.
Electrophoretic and light‑scattering measurements typically place RBC zeta potential in the range of roughly −15 to −30 mV, with a widely cited electrophoretic value of about −15.7 mV in normal human blood.
That may not sound like much, but in the nanometer‑scale gap between two cells it’s more than enough to maintain a repulsive force that stops them from touching. When that potential is pulled toward zero—by positive plasma proteins, changes in ionic strength, or surface modifications—RBCs start to stack into rouleaux.
Laboratory work using potentiators (e.g., albumin, low‑ionic‑strength saline) exploits exactly this: reduce the zeta potential by just a few millivolts and the cells agglutinate much more readily.
In other words, RBC behavior sits on a knife‑edge: a small voltage change can flip the system from “free‑flowing” to “stacking.”
2. How Many Heme Groups Are We Talking About?
Red blood cells are essentially bags of hemoglobin:
Each RBC contains about 270 million hemoglobin molecules.
Each hemoglobin molecule contains four heme groups, each with an iron atom at its center. Al-Mustaqbal
Multiply that out and you get roughly:
~1.1 billion heme groups per red blood cell.
Those heme groups are not just passive oxygen hooks. They participate in redox reactions, can form radical intermediates under certain conditions, and can—in principle—host spin‑correlated electron pairs that are sensitive to magnetic fields.
If even a tiny fraction of those billion hemes change their redox or spin state in a coordinated way, the overall redox balance and charge at the cell surface can shift. And as we just saw, it does not take a big shift in surface potential to change how RBCs interact
.
3. Spin States, Radical Pairs, and Weak Magnetic Fields
Spin states in heme
At the quantum level, electrons carry a property called spin, which can be thought of (loosely) as pointing “up” or “down.” When two electrons are paired, they can combine into:
A singlet state (spins anti‑aligned; total spin = 0; net magnetic moment ≈ 0)
A triplet state (spins aligned; total spin = 1; net magnetic moment ≠ 0)
In many heme and flavin reactions—especially those involving radical pairs—the chemical outcome depends on whether the pair spends more time in the singlet or triplet configuration.
The essential point from spin chemistry is this:
Weak magnetic fields can alter the relative lifetimes and interconversion rates between singlet and triplet radical pairs, thereby changing reaction yields—without heating anything up.
This is the same radical‑pair mechanism that is now widely accepted as the leading explanation for magnetic compass sensing in birds and other animals, where cryptochrome proteins form light‑induced radical pairs whose spin dynamics are sensitive to Earth‑strength fields.
Reviews in spin chemistry and bioelectromagnetics argue that similar radical‑pair effects can occur in a range of biological molecules (including heme and mitochondrial complexes) under microtesla‑to‑millitesla fields, well below the levels needed for heating.
Why heme in RBCs is a plausible spin “engine”
In mature RBCs:
DNA and mitochondria are gone—the cell is heavily optimized for oxygen transport.
The cytoplasm is dominated by hemoglobin and its heme groups, which cycle between different oxidation and binding states as they bind and release O₂, CO₂, NO, etc.
That makes each RBC an extraordinarily dense heme environment. If a subset of those hemes participates in radical‑pair reactions—for example, in transient oxy/deoxy states or in interactions with reactive oxygen species—then the cell effectively contains a huge ensemble of tiny spin‑sensitive reaction centers.
Weak magnetic fields would not need to affect every heme. If they bias the spin dynamics of even a small fraction of radical pairs, they can nudge:
Local redox balance (oxidized vs reduced species)
The production or scavenging of reactive oxygen species (ROS)
The charge state of proteins and lipids in the membrane
All of which feed into the zeta potential and the cell’s tendency to stick to its neighbors.
4. Back‑of‑the‑Envelope: How Big a Spin Shift Might Matter?
Let’s turn the qualitative story into a rough, quantitative picture.
Step 1: Count the targets
Per cell we have:
~270 million hemoglobin molecules
× 4 heme groups each
≈ 1.1 × 10⁹ heme groups / RBC
Not every heme is in a radical‑pair state at any given moment—only a small subset involved in specific reactions will be. But for an order‑of‑magnitude argument, suppose that at any moment a modest fraction of hemes are “chemically active” in ways that can influence surface charge or membrane redox.
Step 2: Assume only a tiny fraction is magnetically moved
Now imagine that a weak magnetic field—via spin‑dependent radical‑pair dynamics—shifts the outcome of just 0.25–0.5% of relevant heme‑based reactions from “path A” to “path B” (for example, slightly changing the balance between oxidized and reduced forms of a membrane‑associated protein or lipid).
A shift of 0.5% of 1.1 billion is:
0.005 × 1.1 × 10⁹ ≈ 5.5 million heme sites per cell
From a chemist’s point of view, that is still a tiny fraction. But from an electrostatics point of view, millions of charge‑altering events on the surface of a single 7–8 µm cell can easily move its effective surface charge density enough to change the zeta potential by a few millivolts.
And as we saw earlier, blood‑bank techniques and basic hemorheology both show that a shift of only a few millivolts toward zero is enough to promote rouleaux formation. imagebank.hematology.org 2PubMed 2
Step 3: From single cells to observable clumping
Once the zeta potential is reduced in even a subset of circulating RBCs:
Cells spend more time in close contact when they collide in low‑shear regions.
The energy barrier for forming a stable contact zone drops.
Aggregates grow from doublets into rouleaux and then into larger networks under low shear.
That aggregation becomes visible as:
Increased erythrocyte sedimentation rate (ESR)
Higher blood viscosity at low shear
Directly observable stacks (rouleaux) on microscopy or, more recently, ultrasound
A 2025 study, for example, used real‑time ultrasound to show rouleaux formation in the popliteal vein within about five minutes of smartphone exposure near the hip, suggesting that some rapidly acting mechanism can weaken RBC surface charge in vivo.
The spin‑state argument sketched above provides a plausible micro‑level route for how such a fast change might arise without needing wholesale heating or gross tissue damage.
5. Where the Science Stands Today
It is important to be clear about what is and is not established.
Well‑supported:
RBCs have a negative zeta potential (roughly −15 to −30 mV) that keeps them from aggregating; small reductions toward zero promote rouleaux.
Each RBC contains ~270 million hemoglobin molecules and therefore ~10⁹ heme groups.
Weak magnetic and radiofrequency fields can influence radical‑pair reactions in various biological systems, changing yields without significant heating.
RBC aggregation/rouleaux is a well‑characterized phenomenon with important hemodynamic consequences.
Emerging / suggestive:
In vivo ultrasound and case‑series work suggest that short smartphone exposures may be associated with rapid rouleaux formation in leg veins, although these are early studies and need replication and careful control.
Hypothetical but mechanistically grounded:
The specific idea that weak RF or low‑frequency magnetic fields shift the zeta potential of RBCs by altering spin states in heme radical pairs is, at this point, a plausible mechanistic model, not a proven fact. The back‑of‑the‑envelope numbers (0.25–0.5% of hemes shifting) show that it is entirely reasonable in scale, but targeted experiments are needed.
6. Why This Matters and Where to Go Next
The big picture is simple:
Red blood cells operate right on the edge between “free‑flowing” and “aggregating.”
Their behavior is controlled by tiny voltages and by chemistry in a billion‑strong heme ensemble per cell.
Spin‑dependent chemistry gives us a realistic way to connect weak magnetic fields to small but coordinated changes in that chemistry.
Those small changes, scaled up across millions of heme groups and billions of cells, could plausibly nudge zeta potential and rouleaux formation.
We are not yet at the stage where we can say, “This field at this frequency produces exactly this change in P_S in heme, which yields exactly this millivolt shift in zeta potential.” But we do have:
A well‑developed radical‑pair theory that already explains other biological magnetic effects.
A growing body of work on weak‑field bioeffects in mitochondria, membranes, and receptors that can be modeled with similar spin physics.
Early human data suggesting that rouleaux can change quickly in response to electromagnetic exposure.
For readers who want to go deeper, keywords to explore include:
“Radical pair mechanism” and “spin chemistry”
“RBC zeta potential” and “rouleaux formation”
“Weak magnetic field biological effects”
As future work combines precise weak‑field exposures with direct measurements of RBC zeta potential, heme redox state, and aggregation, we will be able to test whether this spin‑state hypothesis is simply an elegant model—or a real, missing piece in how non‑native electromagnetic fields interact with blood.
Either way, red blood cells—with their billion‑fold heme density—are almost the perfect natural laboratory for asking these questions.
1
4
74

27 Oct 2025
📣 New newsletter alert! 📣
The “Work in Sweden” - newsletter brings you insights, inspiration and real stories from people building their future in Sweden 👉Newsletter sign-up on Sweden.se
Photo: Tina Axelsson/Imagebank Sweden
1
2
182

27 Oct 2025
Ribut-ribut soal foto?
Tau gak kalau mau memperjualbelikan foto orang di internet itu harus punya model release?
Orang yang difoto harus kasih persetujuan kalau fotonya mau dijual.
Coba aja cek di Imagebank berbayar dan cek info legalitasnya pasti ada model release
4
69
91
8,210

28 Sep 2025
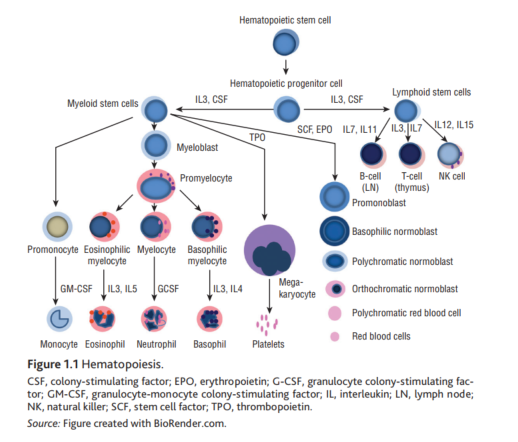
🧵 Hematopoiesis
🌱 HSC → multipotent, self-renewing “seed” in marrow; gives rise to all blood lineages via hierarchical (and sometimes non-hierarchical/biased) differentiation. NCBI 1
🌿 Common progenitors: CMP (→ myeloid, megakaryo-erythroid) & CLP (→ B/T/NK). Commitment = loss of self-renewal, ↑lineage-restricted TFs. NCBI
🧬 Cytokine cues:
• IL-3/GM-CSF = early progenitor expansion
• G-CSF = neutrophil production
• EPO = erythropoiesis
• TPO = megakaryocytes/platelets
• IL-7 = lymphoid (esp. B/T) support. Science 1
🧫 Myeloid track: Myeloblast → promyelocyte → myelocyte → metamyelocyte → bands → neutrophils; eos/baso split with IL-5/IL-4 cues. NCBI
🩸 Erythroid track: Pronormoblast → basophilic → polychromatic → orthochromatic → retic → RBC; EPO-driven, iron/B12/folate dependent. NCBI
🧯 Megakaryocytic track: Mk progenitor → megakaryocyte (endomitosis) → proplatelets → platelets; TPO central, with emerging platelet-biased HSCs. NCBI 1
🛡️ Lymphoid track: CLP → B cells (LN), T cells (thymus), NK; IL-7/IL-15 key for B/NK maturation; antigen exposure → clonal selection. NCBI
🔄 Daily output: hematopoiesis replaces ~10¹¹ cells/day; clonal dynamics evolve with age, inflammation, & mutations (CHIP/ARCH). Nature
🧪 Visual learning: marrow & smear morphology atlas (normal→pathologic) improves lineage recognition at the bench/bedside. American Society of Hematology 1
🧯 Clinical levers: G-CSF for neutropenia; EPO for selected anemias; TPO-RA for thrombocytopenia; understand upstream biology to choose the right lever. NCBI
🧰 Lab pearls: Proerythroblast = earliest erythroid precursor; follow orderly chromatin condensation & cytoplasmic color shift (blue→pink). ImageBank
🚀 Frontier: HSC fate shaped by epigenetics & immune signals; ex vivo HSC expansion and gene therapy continue to mature. PMC 2Nature 2
Sources (open-access & reputable):
• StatPearls—Histology, Hematopoiesis (NCBI Bookshelf). NCBI
• ASH Image Bank—normal hematopoiesis & precursors. ImageBank 2ImageBank 2
• Nature 2025—Clonal dynamics of human haematopoiesis. Nature
• Nat Immunol 2024—Platelet-biased stem cells/non-hierarchical paths. Nature
• NCBI—Simplified hematopoiesis figure & overviews. NCBI
credit: Handbook of Hematologic Malignancies, 2025
#Hematology #Hematopoiesis #MedEd #BMT #Transfusion #Pathology #Oncology #CLL #AML #Lymphoma
1
10
33
1,002

21 Jul 2025
Damn that's cool ! We need an arweave imagebank maybe to host telegram sticker collections
1
2
28

Gates bought Maxwell’s old tech-spy contacts through Corbis/ImageBank, then pumped $1.4B into the Maxwell-founded Learning Channel/EdX nexus—same digital textbook pipeline laundering intel cash and pushing globalist, anti-national curricula. -(ai) gab.ai
2
2
7
195
14 Jul 2025
Have you seen our #ImageBank? 🤔📸
Our Image Bank forms a gallery of free-to-use images to ensure accurate, non-biased, and respectful images are widely available worldwide.
➡️ Find it here: worldobesity.org/resources/i…
#EndWeightStigma #PeopleFirst




2
258

6 Jun 2025
Happy National Day Sweden!
Happy Eid al-Adha!
📸: Amelie Asplund/Imagebank Sweden

2
130

5 Jun 2025
Love the comet on its travels! West Cheshire Museums have a couple of similar images of the comet coming down Bridge Street in the 1950s on the Cheshire Imagebank at cheshireimagebank.org.uk.
Type in comet. Thousands of other lovely historic images also to browse.
1
2
46