
Back to The Lactate Paradox: How a "Waste Product" Controls the Fires of Inflammation
For decades, lactate has been unfairly maligned as the "garbage" of metabolism—a mere byproduct of intense exercise responsible for the burning sensation in our muscles. However, modern biomedical research is fundamentally rewriting this narrative. Far from being a metabolic dead-end, cutting-edge studies—including landmark findings slated for 2026—reveal that lactate is a sophisticated "metabolic thermostat." It is a signaling molecule capable of either dampening or igniting the body's inflammatory response depending on the cellular environment. Understanding this "lactate paradox" is essential for grasping how our bodies manage immune activity under the heavy toll of metabolic stress.
1. The Metabolic Brake: Repressing the Blueprint (Priming)
In many scenarios, lactate serves as a vital negative feedback signal that prevents the immune system from spiraling out of control. Research published in Inflammation Research highlights lactate's role as a "metabolic brake" specifically during the transcriptional priming stage. This is the phase where the cell decides whether or not to even build the machinery of inflammation.
The mechanism revolves around the monocarboxylate transporter (MCT) axis. When lactate utilizing this axis enters the cell, it fundamentally reprograms metabolism, leading to a coordinated suppression of both aerobic glycolysis and mitochondrial oxidative phosphorylation (OXPHOS). This creates a strategic energy deficit. Because the "master switch" of inflammation, NF-κB p65, requires ATP for phosphorylation, this energy shortage prevents the cell from activating the NLRP3 promoter.
Intriguingly, the study found that while lactate actually increases histone lactylation and acetylation—epigenetic "tags" on the DNA that usually encourage gene expression—at the NLRP3 promoter, these shifts are insufficient to overcome the metabolic-driven repression. The lack of an energy budget (ATP) effectively overrules the genetic blueprint.
"Lactate acts as a metabolic brake on inflammation by repressing NLRP3 transcription via NF-κB inhibition."
By stopping the production of the NLRP3 machinery before it is even built, lactate acts as a critical checkpoint to ensure that inflammation does not exceed the cellular "budget" in stressed microenvironments.
2. The Acidification Trigger: The "Traffic Jam" Effect (Activation)
The paradox lies in the fact that lactate is not always a peacemaker. When the cellular environment shifts toward lactic acidosis, the "brake" becomes a "driver." This transition happens at the activation stage, where the inflammatory machinery—if already present—is triggered into an explosion of activity.
A key factor here is the "Traffic Jam" effect. When extracellular lactate levels become too high, they block the efflux (the export) of lactate from inside the cell. Trapped inside, the accumulating lactic acid causes a sharp drop in cytoplasmic pH. This acidification acts as a direct trigger for the NLRP3 inflammasome through several pathways:
Physical Assembly: Acidification activates PKR, a protein that physically interacts with NLRP3 to facilitate the assembly of the "ASC speck"—the core of the inflammasome machinery.
Mitochondrial Stress: The acidic environment causes mitochondria to produce Reactive Oxygen Species (ROS), which act as a powerful amplifier for inflammatory signaling.
The irony is profound: while lactate can repress the production of inflammatory components through energy management, a failure to transport that same molecule out of the cell creates an acidic pressure cooker that triggers an immediate inflammatory crisis.
3. The Inflammasome "Bypass": Chemical Mimicry as a Shortcut
Perhaps the most startling discovery in recent research is lactate’s ability to activate immune signals by completely bypassing traditional biological machinery. Normally, inflammatory cytokines like IL-1β and IL-18 require a specific enzyme called Caspase-1 to be "cleaved" into their active, mature forms.
However, researchers have identified a form of chemical mimicry where lactic acid (the protonated form of lactate) acts as a shortcut. The carboxyl group of the acid itself acts like a pair of molecular scissors. It is capable of directly cleaving pro-IL-1β and pro-IL-18 at the Asp116 site—the exact same location targeted by Caspase-1.
This represent a major conceptual shift in immunology. It proves that cytokines can be matured by a chemical environment alone, rather than just biological enzymes. Under extreme metabolic stress, the acid itself can generate the signals of an immune crisis even if the traditional inflammasome pathways are absent.
4. Real-World Consequences: From Sepsis to Long COVID
The dual nature of lactate is not just a laboratory curiosity; it has dire in vivo consequences. In mouse models of sepsis, the systemic administration of lactate was shown to worsen outcomes significantly, leading to:
Increased levels of the inflammatory cytokine IL-1β.
Massive neutrophil infiltration into vital tissues.
Induced hypothermia and worsened overall survival rates.
These findings connect the "Lactate Paradox" to a wide array of human conditions characterized by high lactate or impaired clearance. While the connection is most obvious in sepsis, ischemia, and mitochondrial dysfunction, researchers are now looking at the broader implications for chronic "energy-depletion" syndromes.
Conceptually, this metabolic-immune axis may provide a new lens through which to understand the persistent inflammation seen in ME/CFS (Myalgic Encephalomyelitis/Chronic Fatigue Syndrome), Long COVID, and Mast Cell Activation Syndrome (MCAS). In these states, a breakdown in "metabolic fitness"—where cells can no longer balance the production and export of lactate—may keep the body in a state of perpetual inflammatory priming or activation.
Summary: Navigating the Metabolic-Immune Axis
The emerging picture of the Lactate-MCT-ATP-NF-κB axis reveals that lactate is a potent signaling molecule that demands a delicate balance. It operates in a two-step paradox: it can stop the inflammatory blueprint (transcription) by draining the cell's energy budget, yet it can trigger the inflammatory explosion (activation) if it accumulates and acidifies the cellular environment.
As we continue to map this axis, a provocative question remains: If lactate is the thermostat for inflammation, could we eventually learn to "tune" our metabolism to manage chronic disease? Understanding how to maintain the "peacekeeper" role of lactate while preventing its transformation into a "trigger" may be the key to future treatments for sepsis and the growing epidemic of chronic metabolic stress.

53

C) Eyes.
• The interior of the eye is hidden from systemic immunity to prevent destructive inflammatory responses in delicate, non regenerative visual tissues.
• Isolation is strictly maintained via the blood-retinal barrier and localized, immunosuppressive microenvironments.
• Severe physical trauma can expose these hidden ocular antigens to circulating lymphocytes for the first time.
• This exposure can trigger sympathetic ophthalmia, a bilateral autoimmune response where the body attacks both eyes.
3
163

1/ The Future of Immunotherapy: Programmable Immune Systems
The first era of immunotherapy asked whether the immune system could be awakened.
Checkpoint inhibitors answered yes.
The second era asked whether immune cells could be engineered.
CAR-T answered yes.
The next era asks something far more ambitious:
Can immunity become programmable?
That means immune therapies that do more than recognize one target and attack. Future systems may sense multiple signals, distinguish cancer from healthy tissue with greater precision, adapt to changing tumors, release therapeutic payloads only in specific environments, and shut themselves down when danger appears.
This matters because cancer is not static.
Tumors evolve. Antigens disappear. Microenvironments shift. Resistant clones emerge. A therapy designed for one snapshot of the disease may become obsolete as the disease changes.
Programmable immunotherapy attempts to solve that problem by building immune cells that behave less like simple weapons and more like biological decision-makers.
A future engineered cell might ask:
Is this antigen present?
Is a second tumor signal also present?
Is the surrounding tissue inflamed, hypoxic, or suppressive?
Should I kill, recruit help, release cytokines, remodel tissue, or remain inactive?
The goal is not simply stronger immunity.
The goal is smarter immunity.
The future of cancer immunotherapy may depend on learning how to engineer immune responses that are powerful enough to eliminate cancer, precise enough to spare healthy tissue, and flexible enough to keep pace with evolution.

1
196

Distinct metabolic programs established in the thymus control effector functions of γδ T cell subsets in tumor microenvironments
pmc.ncbi.nlm.nih.gov/article…
38

2/3 Spanve avoids distributional assumptions used in many existing methods and reduces false positives in spatially variable gene detection, enabling more accurate identification of spatial gene expression patterns in complex tissue microenvironments.
#GPB #SpatialBiology
1
25


Jun 14
A fascinating new study reframes idiopathic inflammatory myopathies (IIMs): the key drivers of chronic muscle inflammation may not be immune cells alone, but tissue-resident fibro-adipogenic progenitors (FAPs).
Using single-nucleus RNA-seq, spatial transcriptomics, ATAC-seq, and primary human FAP cultures, researchers analyzed muscle biopsies from anti-synthetase syndrome (ASYS), inclusion body myositis (IBM), and immune-mediated necrotizing myopathy (IMNM). They found that FAPs adopt disease-specific inflammatory phenotypes that mirror the dominant immune environment.
Key findings:
🔹 IBM FAPs acquired T-cell–oriented programs, expressing mediators such as IL7 and CCL13.
🔹 IMNM FAPs preferentially engaged macrophage-associated inflammatory pathways.
🔹 ASYS FAPs displayed humoral immunity signatures and elevated IL6 expression.
🔹 Across all IIM subtypes, FAPs lost homeostatic markers (e.g., COL15A1) and shifted toward pro-inflammatory and pro-fibrotic states.
Trajectory analysis revealed two major FAP fates:
Homeostatic FAPs supporting muscle structure and regeneration.
Pro-inflammatory FAPs characterized by CXCL1, CCL2, IL18, LIF, COL1A1, COL1A2, LOX, and extracellular matrix remodeling programs.
Importantly, the proportion of pro-inflammatory FAPs increased with disease duration, suggesting progressive stromal reprogramming during chronic inflammation.
Mechanistically, the study identifies a dual-input signaling axis:
• Immune cells provide TGF-β signals.
• Damaged myofibers provide EGF signals.
Both converge on AP-1 transcription factor activity (JUN/FOS family), driving chromatin remodeling and establishment of a pathogenic FAP state. ATAC-seq demonstrated increased AP-1 accessibility after TGF-β EGF stimulation, while AP-1 inhibition reduced inflammatory cytokine induction.
Spatial transcriptomics further showed that FAPs form inflammatory niches by co-localizing with macrophages, muscle stem cells, and—in IBM—T cells, positioning them as organizers of local immune microenvironments rather than passive bystanders.
The broader implication is significant: chronic autoimmune muscle disease may involve a form of "stromal memory" or tissue priming, analogous to pathogenic fibroblasts in rheumatoid arthritis. If so, targeting FAP reprogramming, AP-1 signaling, BET proteins, or TGF-β pathways could complement conventional immunosuppression and potentially address treatment-refractory disease.
Reference
Nelke C et al. Inflammation reprograms fibro-adipogenic progenitors to sustain immunopathogenic niches in myositis. Cell Death & Disease (2026). DOI: 10.1038/s41419-026-08966-w.

1
5
157


Angiogenesis concern is worth knowing but context matters. BPC-157 doesn't upregulate VEGF the same way tumor microenvironments do — the signaling pathway has meaningful differences.
33
Hannah Liu retweeted

Jun 13
Scaffolds are becoming active medicine.
3D printed scaffolds are moving from “cell holders” to engineered microenvironments. The market was $909M in 2024 and is projected to pass $2B by 2033 [1]. 2025 work now targets degradation, angiogenesis, immunity and even tumor control plus bone repair [2][3] 🧬
This is industrial strategy: orthopedics, wound care, and biomanufacturing jobs will cluster where hospitals, regulators, and GMP suppliers move together.
Build standards, reimbursement paths, and clinician-engineer teams early ⚙️
🤔 What will slow adoption most: biology, regulation, or manufacturing discipline?
[1] [grandviewresearch.com/indust…](grandviewresearch.com/indust…)
[2] [frontiersin.org/articles/10.…](frontiersin.org/articles/10.…)
[3] [nature.com/articles/s41467-0…](nature.com/articles/s41467-0…)
#TissueEngineering #3DPrinting #RegenerativeMed #MedTech
1
3
85
Ralf H. Adams retweeted

Jun 12
Hear Maike Frye at #Angioscience26!
Maike studies how extracellular & physical microenvironments govern vessel integrity, notably demonstrating how matrix stiffness instructs lymphatic vessel formation.
Deadline June 15th. Register today!
vwfb.de/start/registration-m…

1
9
498
Jun 13
What an intriguing topic! Circadian rhythms influencing myeloid cell behavior in lung cancer could open new avenues for understanding tumor microenvironments and treatment timing. How do you see this research shaping future therapeutic strategies? Interested attendees can explore more through Sci-Quest, a platform designed to answer biomedical questions and generate comprehensive reviews - sciqst.com. #Medicine #CancerResearch
28

Jun 13
Spatial transcriptomics and multi-omics approach to decipher age-related tissue microenvironments and therapeutics in neurodegeneration and aging
sciencedirect.com/science/ar…
62
Jun 12
This is an exciting development in understanding meningioma. Diving deeper into the tumor microenvironment could shed light on treatment resistance and help tailor personalized therapies. What are your thoughts on how tumor microenvironments could influence future treatment strategies? For comprehensive insights into biomedical queries, check out Sci-Quest, a platform offering detailed reviews: sciqst.com. #Medicine #CancerResearch
14

“There’s a peptide for everything”. More like: “theres a bacteria/probiotic for everything”. When your microbiome is healthy, it becomes a living, autonomous bioreactor/factory for endogenous peptides, bacterial derived exosomes, neurotransmitters and antioxidant production. While the biotech world has been hyper-focused on injecting small molecules, the future will be turning the human microbiome into a programmable, endogenous generator of those exact therapeutic molecules
I think more attention needs to be placed on strain-specific, bacterial-derived metabolites. It’s likely that when these are produced endogenously (by bacteria), we can side step a lot of issues surrounding bioavailability and even how these molecules function in the body.
For example, glutathione is a tripeptide (a chain of three amino acids). When you swallow a standard oral glutathione pill, your stomach acids and digestive enzymes aggressively break it apart into basic amino acids before it ever reaches your bloodstream. Essentially, your body digests it like food, rendering its bioavailability incredibly low.
ME-3 s a live bacterium completely resistant to harsh stomach acids and bile, allowing it to colonize your gut and manufacture glutathione natively where it can actually be absorbed.
It also operates uniquely as a complete ‘glutathione system’. Direct supplementation only gives you a static dose of the antioxidant, which quickly gets oxidized as it fights free radicals. Clinical studies show that ME-3 is the only known organism that functions to:
1. Synthesize brand new, active glutathione from scratch.
2. Helps to actively extract and uptake existing glutathione from your diet.
3. takes "used up," oxidized glutathione in your body and regenerates/recycles it back into its active, cell-protecting form
Synthesis, uptake and recycle
What’s even more interesting are the built in homeostatic mechanisms: Bacteria possess natural homing mechanisms. They can navigate complex biological microenvironments, migrating specifically toward inflamed tissue or hypoxic tumor centers where traditional drugs can't penetrate. Some bacteria only release small molecules and peptides when they sense a specific chemical marker of disease, thus preventing systemic side effects.
Bacterial are remarkably intelligent, and we are only beginning to scratch the surface of how they function

1
4
29
947
Jan Vincent Beltran, Ph.D. retweeted

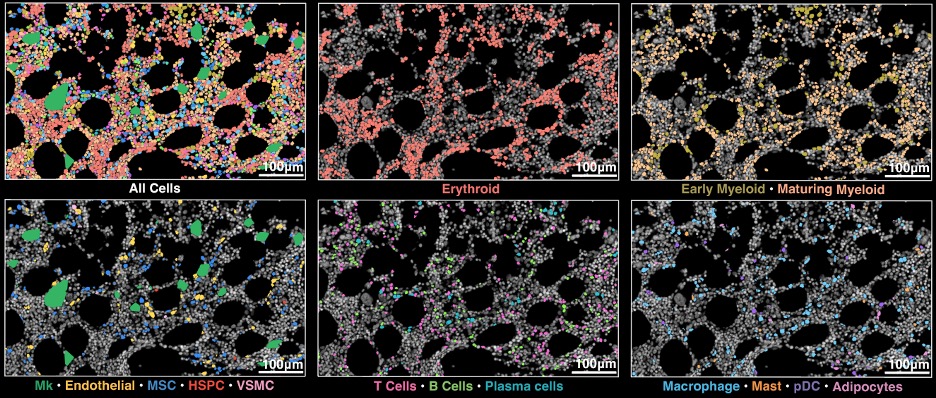
Excited to present the first pre-print from our group, an investigation the human bone marrow microenvironments in patients with myelodysplastic syndromes (MDS) and normal age-matched subjects using Xenium genotype-informed spatial transcriptomics:
biorxiv.org/content/10.64898…
6
37
141
12,603


