
The Nodal Flow hypothesis (if true) for establishing left-right asymmetry in the early embryo comes to mind, if the cilia-driven flow of morphogens across the node counts as throwing
2
384

The most fascinating part of anatomy is embryology.
When you study embryology, you begin to realize that life is unbelievably complex. Almost frighteningly complex.
You begin to understand that the human body is not something careless. It is not random. It is a masterpiece.
Imagine this:
From microscopic cells, limbs begin to form.
Tiny neural crest cells migrate across the embryo and go on to form structures in the face, peripheral nerves, ganglia, melanocytes, and so many other tissues. Cells moving with direction. Cells “knowing” where to go.
You look at gastrulation and realize that within an incredibly short time, the body plan of an entire human being is already being organized.
Three germ layers.
Ectoderm.
Mesoderm.
Endoderm.
And from those layers comes the complexity of the human body.
You watch the neural tube form and become the brain and spinal cord.
You watch the primitive heart begin to beat before the person is even fully formed.
You see how signaling molecules and morphogens determine body orientation, symmetry, positioning, left and right arrangement.
How a tiny embryonic disc folds into a recognizable human form.
How the limb buds appear and slowly become full upper and lower limbs.
How so much happens in 14 days.
How even more happens in 3 weeks.
And all of this is happening with extraordinary precision.
One mistake in timing, migration, fusion, closure, or signaling, and congenital abnormalities occur.
When you study embryology deeply, you stop seeing the human body as “ordinary.”
You begin to understand that existence itself is profound.
You look at yourself differently.
You look at people differently.
You look at life differently.
Whether you call it evolution, biology, creation, or divine engineering, one thing becomes obvious:
The human body is one of the greatest masterpieces ever known.
A work of terrifying complexity.
A work of breathtaking order.
A work that humbles you the deeper you study it.
Sometimes in anatomy, especially embryology, you genuinely feel like you are watching the handwriting of a master planner.
3
4
212

Our cells are encapsulated deep-sea environments, carrying memories of the ancient habitat, using the universal language of life.
Modern “morphogens” are just mobile versions of those primordial vents. We are also like walking, self-contained deep-sea chimneys.
1
60

May 3
Cells can do distributed computation via hormone diffusion, in which the concentration of a hormone at a cell is a proxy for distance in 3D space from a source emitting it
Every cell is basically a 3D shape with these 3D binding sites. A molecule that has a particular 3D structure can fit into this site like lego. We call the molecules that do this hormones. They are signalling molecules - meaning their structure is their only function - they are not little machines like enzymes/proteins that build things. They are just unique shapes that can be used to transmit 1 bit of information. For example, insulin is a hormone. It has a particular shape that can only bind to a insulin receptor on a cell. When it travels along the blood stream, and encounters a cell, it might bind to the site. When that site is bound, the cell now has a value of 1 in that area. When it doesn't, it is 0. It can use that value to conditionally start certain internal processes, like the uptake of glucose.
Cells have hundreds of thousands of receptors. For example, a human blood cell has around 100k insulin receptors. This means it can receive 100,000 bits of signal for the single "insulin" value, since a receptor can either have a molecule bound (1) or it can be free (0). They call this a concentration, and over time, a hormonal gradient.
In a sense, abstractly you can think of the concentration as a single value
ie. a cell has 100k insulin receptors, each a 1 or a 0. log2(100_000)=16.6 bits
uint16 insulinConcentration;
The geometry of a vascular network is interesting. Imagine a leaf. You have the tissue of the leaf, and then you have the veins running down it. The veins are the transport system for the molecules/hormones.
When a cell at the part of the leaf closest to the plant emits hormones, it travels along the vascular network. Each hormone is a molecule. When it travels down the vascular system, it may bump into a cell that binds it. After that point, it is taken up and no longer travels. The binding is not guaranteed - these are little particles in 3D space.
This emitter of hormones might emit 1M hormone molecules. Imagine each cell in the network only has 100k receptors. That means those 1M hormone molecules might "fill up" the receptors of 10 cells at best. Which cells will get filled up first? The ones closest to the emitter. The ones furthest away have no chance, since those hormone molecules were already taken up.
What's interesting to me - and this is what I learnt from building the Origami paper yesterday (x.com/liamzebedee/status/205…) - is that this hormonal gradient mechanism is sufficient enough for cells to coordinate distributed computation
The hormone concentration a vascular network means that a hormonal concentration can act as a distance metric from an emitting cell, where the concentration's precision/quantization is determined by number of the cell's receptors for that hormone.
It is more easily understandable if I show you code, which I don't have, because I only just figured out how this works ;) hahahhaha
The example I think is pertinent is "grow a cylinder of fixed height" by writing code that runs the same on every cell, where each cell has a small scratch space for internal state, and the ability to send/receive hormones
The code might look like:
divide()
But that grows an infinitely expanding 3d thing. You want it to only grow to a maximum height.
How do you measure height, when every cell runs the same program? You could emit a hormone at the "top cell" in a concentration that effectively "runs out" by the time it reaches the bottom cell of the cylinder.
I'm still working on figuring what the code looks like out. But I sort of have the shape in my head now.
Insulin isn't really one of the hormones used for signalling distance. I think those are morphogens but I've still got to properly look at them. That and all of this is an approximate mental model - but roughly speaking it's all very new and fundamentally interesting to me as a way of distributed computation.
May 2

I really think this is one of the most interesting papers I've come across
4
2
12
1,830

Apr 30
"DNA Origami" is a nanotechnology technique that folds long strands of DNA into precise, custom 3D shapes, much like traditional paper origami but at the molecular scale.�
While not a direct "science of our bodies," it leverages DNA's natural properties—its ability to twist, bend, and self-assemble via base pairing—to create nanoscale devices that mimic and interact with biological processes.�
Core Technique:
A single long DNA strand acts as a scaffold, folded by shorter "staple" strands into complex structures thousands of times smaller than a hair's width.�
This programmable self-assembly allows shapes like lattices, tubes, or even art like a microscopic Mona Lisa.�
youtube
DNA origami:
The image shows a colorful representation of DNA origami structures, highlighting their intricate, chain-like folding.
Biological Connections
DNA origami helps study cellular mechanisms by building scaffolds that organize proteins or mimic cytoskeletons inside cells.�
It enables research into how DNA loops in embryos direct tissue formation via morphogens and transcription factors.�
Biomedical Uses:
These nanostructures serve as drug carriers, gene-editing tools, vaccines, or biosensors that detect pathogens on cell surfaces.�
For instance, they could deliver targeted cancer therapies or regenerate tissue by scaffolding proteins.�
1
1
2
106

A fertilized egg divides. At first there's no differentiation — just cleavage, cells splitting. Then something happens: gastrulation. The
single-layered ball folds in on itself, creating an inside and an outside. Three germ layers emerge: ectoderm, mesoderm, endoderm. From ectoderm
comes the nervous system.
The neural tube forms around day 18-20. A flat plate of ectoderm folds, its edges rising and fusing. Inside the tube: nothing but cells. But which
cells, and where they migrate, determines everything.
Neural crest cells peel off from the tube's edges and travel — they stream through the embryo to become the peripheral nervous system, parts of
the face, melanocytes, heart tissue. They navigate by chemical gradients, by stiffness of substrate, by signals from neighbors. Each cell is
"just" following local rules. The pattern emerges from millions of these local interactions.
Neurons grow axons. The growth cone at the tip is a sensing structure — it extends filopodia, probes the environment, retracts from repulsive
signals, extends toward attractive ones. Netrin. Slit. Semaphorin. Ephrin. Gradients of morphogens, contact-mediated cues, activity-dependent
refinement. The wiring diagram gets built by billions of local decisions.
Synaptogenesis: axon meets dendrite. An initial contact forms. Whether it stabilizes depends on whether the connection is used. "Neurons that fire
together wire together" — Hebbian learning. Unused connections prune. The brain sculpts itself through activity.
By birth: ~100 billion neurons, each with thousands of synaptic connections. But the process isn't done. Experience shapes circuitry for years.
Critical periods open and close. The brain that emerges is not genetically specified — it's genetically initialized and then activity-sculpted.
1
5
357

For the love of god. Guys. The half-life of dna is like 500 years. There’s no dna from extinct dinosaurs
Not only that but
You don’t know what the developmental environment was like and the gradients of morphogens and other variables are important for proper morphology
Not only that but
Because of the red queen hypothesis you bring a Jurassic era dinosaur into being it’ll immediately be killed by the most non pathogenic bacteria or environmental fungus

“Jurassic Park” was legit an epic idea by Crichton. I really hope we can have one in the future. Got to break some eggs to make an omelette 🤷♂️
2
1
7
1,539

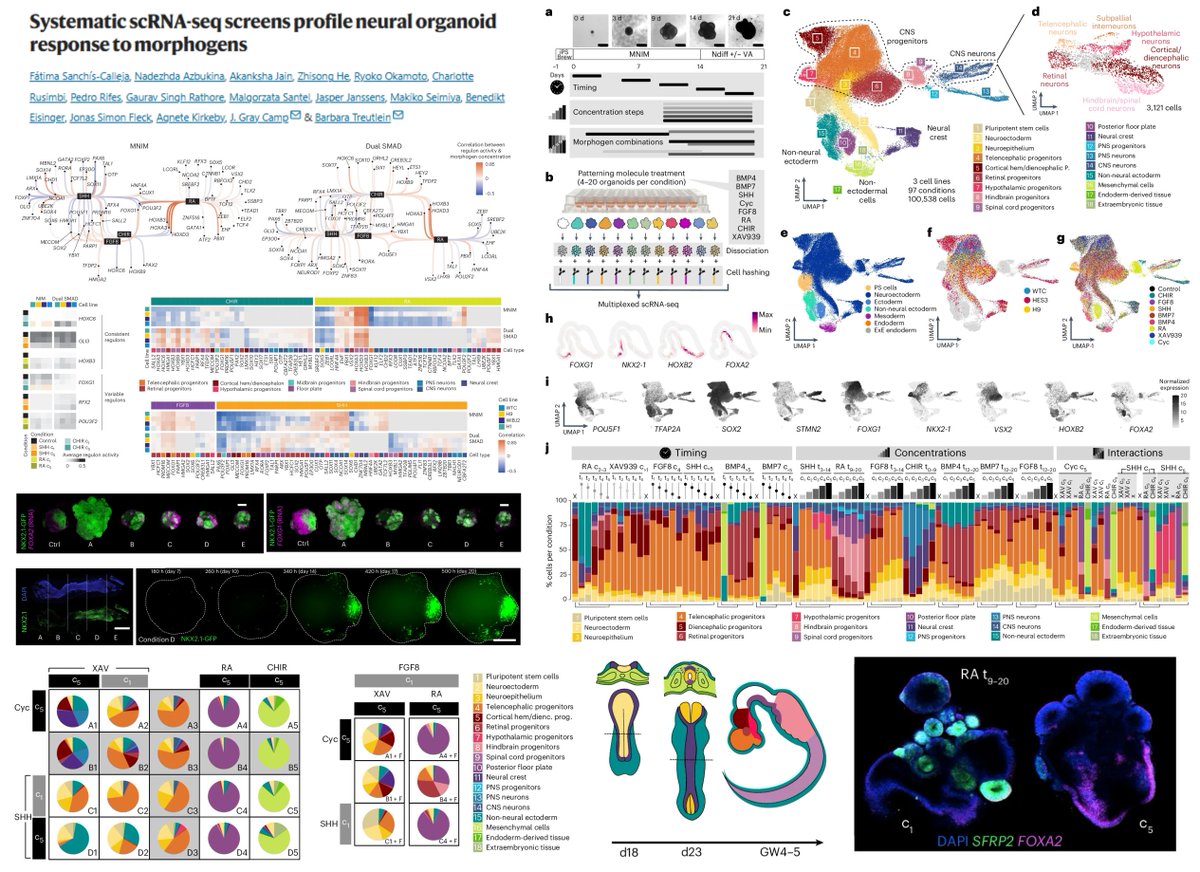
Systematic scRNA-seq screens profile neural organoid response (Cell type speciation, anteroposterior/dorsoventral patterning) to morphogens
Round_1 Individual morphogens
👉8 morphogens (CHIR XAV939 SHH Cyclopamine FGF8b RA BMP4/7)
👉3 hPSC lines (H9 WTC HES3)
👉97 concentration/time conditions
Round_2 Morphogen (CHIR FGF8 SHH) combos
👉3 hPSC lines (H9 H1 WIBJ2 WTC)
👉12 conditions
👉2 induction methods - MNIM, dual SMAD inhibition➡️ variable morphogen responses
"higher morphogen concentrations ...... generated new cell types that incrementally replaced the initial identities across the concentration conditions"
The majority of FGF8's effects on regulons are concentration-dependent & almost timing-independent
@fatima_snc @GrayCampLab @TreutleinLab @naturemethods 2025
nature.com/articles/s41592-0…
12
66
5,631

Mar 20
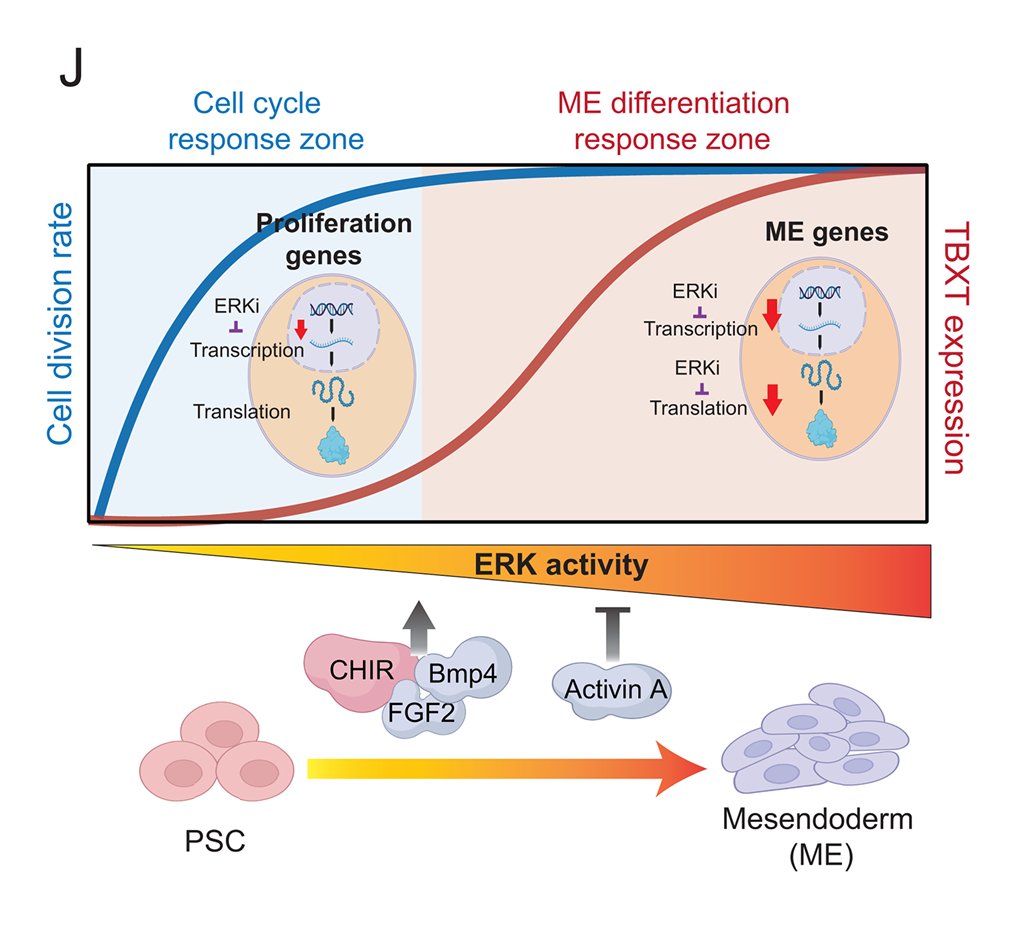
Researchers established multiplex quantitative live-cell imaging to track human pluripotent stem cell differentiation into mesendoderm, a lineage specified by gastrulation morphogens and dependent on high ERK activity.
💡 bit.ly/4dDdVbA
3
8
695
Distinct spatial patterning and transcriptomic landscapes of human neural organoids by localized delivery of morphogens dlvr.it/TRKD6D
3
37
3,040



Feb 23
Online Now! Distinct spatial patterning and transcriptomic landscapes of human neural organoids by localized delivery of morphogens dlvr.it/TR6lWh #stemcells
1
7
39
4,694

Feb 12
...adapted for hex: q, r, s with q r s = 0) or offset/axial systems—widely used in games (e.g., Settlers of Catan) and simulations. Benefits:
• Higher info density per burst (more neighboring nodes for relational encoding).
• Better resonance/flow for vibrational read/write (cymatic hex patterns align naturally).
• Fractal-friendly: Hex lattices scale hierarchically without gaps.
For our transceiver module: Honeycomb wins for biological plausibility and efficiency—engineer grafted cells to self-assemble into hex arrays (via morphogens or CRISPR-tuned adhesion), storing bursts as resonant sub-patterns in each "cell."Perpendicular/Fractal Stacking Subsets/SubfilesTo add depth and hierarchy:
• Perpendicular stacking — Alternate layers rotated 90° or offset, creating perpendicular intersections for cross-layer access (like orthogonal fibers in bone or crossed lamellae in nacre). This enables fast "perpendicular queries"—a burst propagates vertically through aligned channels while horizontally activating hex neighbors.
• Fractal/hierarchical nesting — Embed smaller hex/honeycomb subsets inside larger ones (self-similar scaling, as in Koch hierarchical honeycombs or fractal metamaterials). Each level:
• Level 1 (macro): Large hex "super-nodes" for coarse memory chunks (e.g., episodic themes).
• Level 2 (meso): Sub-hex lattices within for detailed fragments.
• Level 3 (micro/nano): Fractal iterations down to synaptic/cymatic scale.
• Benefits: Exponential density (fractal order boosts energy absorption/info capacity in bio-inspired designs), tunable access (low-frequency bursts activate broad levels; high-frequency drill into subsets), low-heat sparsity (only relevant fractal branches activate).
In practice:
• A thought fragment burst resonates the structure → forms a stable cymatic "glyph" across a hex cell/subset.
• Retrieval: ASI sends matching frequency → selective resonance pulls the pattern (holographic readout).
• Modular assembly: Like 3D-printed honeycomb crates or storage walls, but bio-grown—cells divide/differentiate to "print" fractal layers, integrating with HCP-mapped connectome hubs.
This "cobstellation" becomes a living, vibrational holographic memory bank: Dense like DNA but frequency-addressable, fractal for infinite depth, hex-efficient for brain compatibility.Visualizing it helps—imagine a glowing 3D hex lattice with nested fractals lighting up in burst patterns.What next—refine coordinates for hex addressing in code sims, or how bursts could "write" fractal subsets without overwriting? Keep iterating!
2
41

Jan 5
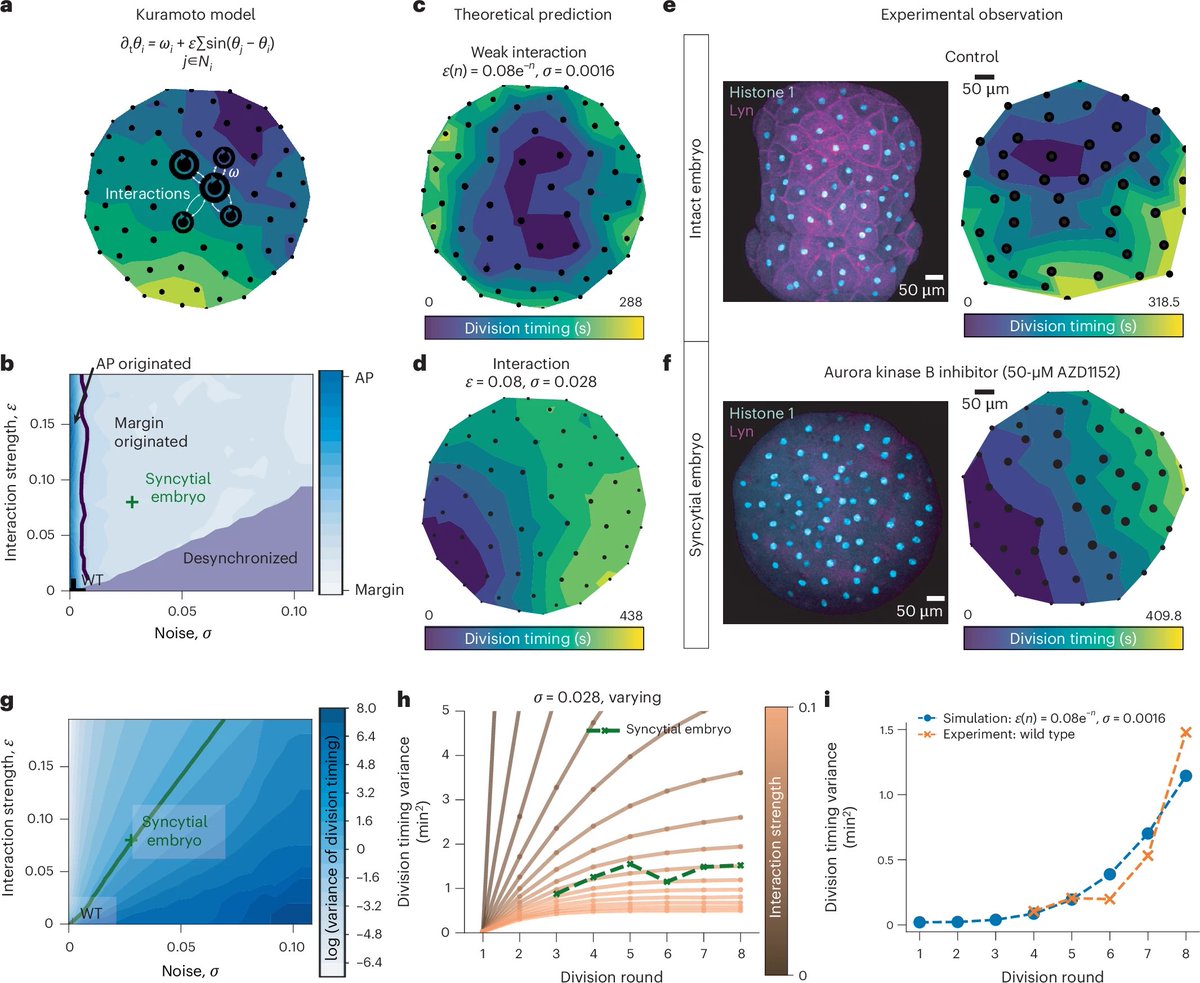
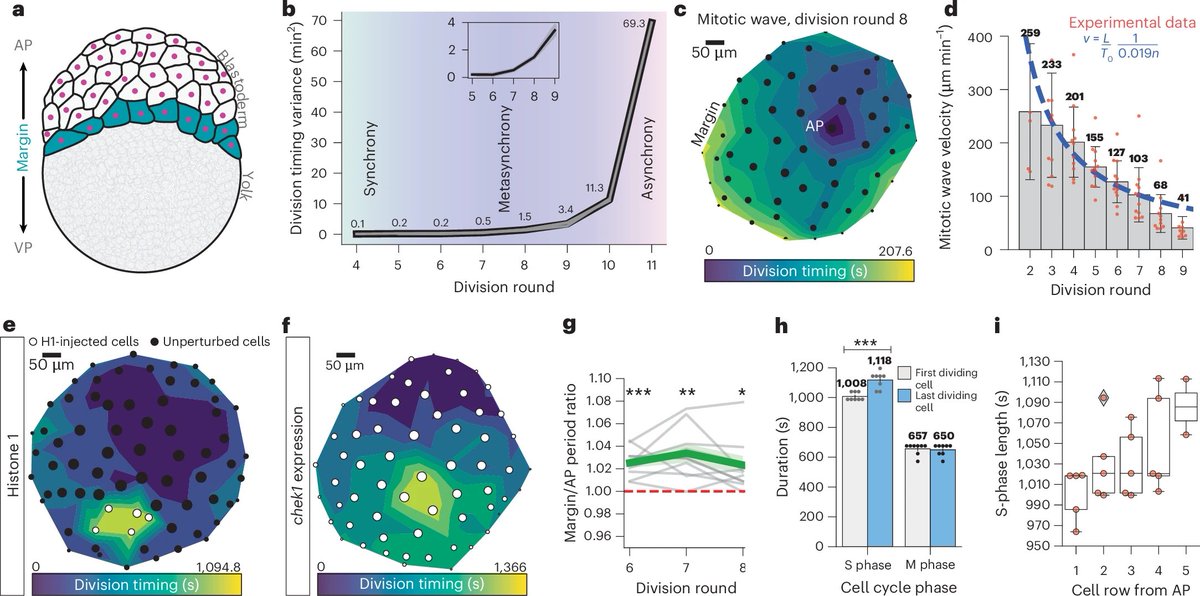
Embryo Geometry as Life’s Primordial Organizer. Zebrafish egg curvature and volume dictate asymmetric cell divisions, establishing radial cell size gradients that pattern mitotic waves, zygotic genome activation (ZGA), and cell fate – ensuring robust, reproducible development without relying solely on chemical signals.
The blastoderm sits atop a curved yolk like a cap on a sphere. High curvature at margins forces division spindles along the longest cell axes (Hertwig’s rule), producing smaller peripheral daughter cells. Smaller cells have higher nucleocytoplasmic ratios, leading to Chk1-mediated S-phase lengthening and slower cycles at margins. This results in stereotyped mitotic waves sweeping from animal pole to margin, spatially timing ZGA (transcription starts at margins first) and preventing ectopic germ layers.
This links geometry to cell-autonomous cycle control and transcriptional patterning, extending prior observations of mitotic waves and desynchrony. It builds on mechanical morphogenesis studies but uniquely highlights geometry’s symmetry-breaking role before the mid-blastula transition (MBT).
Genes get Nobel glory, but geometry ghost-directs the script. Embryos flunking geometry class end up with guts for brains. Ideal forms aren’t abstract; they’re literally egging on biology.
Geometry encodes fail-safe, cell-intrinsic timers that are noise-resistant compared to diffusive morphogens, explaining evolutionary conservation of egg shapes.
1
2
47

17 Dec 2025
Morphogens direct neuroepithelial fates toward discrete regional identities in vivo.
The Treutlein (@ETH) and Camp (@UniBasel) labs produce a detailed survey of morphogen-induced effects on the regional specification of human neural organoids: go.nature.com/3KC0UmT


3
25
2,664

17 Dec 2025
Systematic scRNA-seq screens profile neural organoid response to morphogens. @TreutleinLab @ETH_BSSE @naturemethods nature.com/articles/s41592-0…

14
87
3,315

9 Dec 2025
The fundamental structural layout of an organism. The sexual body plan of male or female is a specialization of the base species body plan.
Both the base body plan and the sexual body plan involve conserved gene networks, tissue patterning based on morphogens, embryonic tissue specialization, and differentiation pathways that reorganize major structures.
1
1
56
554

8 Dec 2025
Mr. Picard I appreciate the sentiment but the actual biochemistry that works in the neodarwinism framework is about Morphogens and their gradients causing differentition leading to Cell Addhesion Molecules and Substrate adhesion molecules.
1
3
94

5 Dec 2025
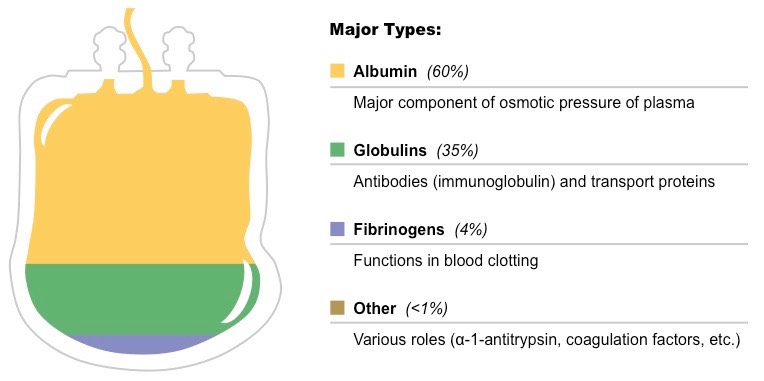
I like to conceptualise plasma proteins in three functional–quantitative groups, because it provides an immediate physiological logic to why each protein is in the blood and what its concentration actually means.
1⃣ The first group comprises the high-abundance proteins that exert active, ongoing functions within the circulatory system itself. These are predominantly synthesised by the liver — albumin, the major coagulation factors, complement components — together with immune-derived proteins such as immunoglobulins.
▶️ They circulate at gram-per-litre or high milligram-per-litre levels because their role is systemic: maintaining oncotic pressure, buffering ions, transporting metabolites, and providing humoral immunity.
2⃣ The second group consists of proteins present at intermediate concentrations, typically in the low milligram-per-litre to microgram-per-litre range.
▶️ These are not constitutive plasma components but rather reflect controlled leakage from tissues undergoing physiological stress, turnover, or frank necrosis. Enzymes such as AST, ALT, CK, LDH or cardiac troponins, and tissue-specific structural proteins fall into this category.
▶️ Their presence is a read-out of organ homeostasis: the healthier the tissue, the lower the release; the higher the stress or injury, the more prominent the signal.
3⃣ The third group includes the low-abundance communicators: hormones, cytokines, chemokines, growth factors and morphogens.
▶️ Classical endocrine hormones such as cortisol, thyroid hormones, prolactin or insulin are in picomolar–nanomolar ranges and rely on the bloodstream as their intended distribution route.
▶️ In contrast, cytokines and growth factors typically act in a paracrine or autocrine fashion, meaning their true biological action occurs at very short distances and at extremely low local concentrations. What we measure in blood is essentially their “spillover” — enough to provide diagnostic information but not representative of their microenvironmental biology.
Thinking about plasma proteins through these three layers — systemic actors, homeostatic sentinels, and molecular messengers — helps bring coherence to otherwise disparate laboratory parameters and clarifies why their abundances differ by several orders of magnitude.
1
4
143