Jun 12
Senescence Is Not Always Bad: It Builds the Brain's Barriers
For years, cellular senescence has been viewed primarily as a hallmark of aging, neurodegeneration, and tissue dysfunction.
But a new Cell study challenges that paradigm.
The authors discovered that senescent cells are an essential developmental program required for formation of the brain's protective barriers.
Rather than representing pathological aging, specific senescent cell states emerge during embryogenesis and actively guide:
🧠 Blood-brain barrier (BBB) development
🧠 Blood–cerebrospinal fluid (CSF) barrier formation
🧠 Brain vascular patterning
🧠 Choroid plexus maturation.
Three brain cell populations become senescent
Using single-cell transcriptomics, lineage tracing, and genetic ablation, investigators identified p21⁺ senescent populations in:
🟡 Choroid plexus epithelial (ChPe) cells
🔵 Vascular endothelial cells (VECs)
🟢 Brain-resident macrophages.
All three populations displayed hallmark features of senescence:
✅ p21 expression
✅ Cell-cycle arrest
✅ Senescence-associated transcriptional programs
✅ Secretory signaling phenotypes (SASP).
However, they behaved very differently.
Two forms of developmental senescence emerge
The study uncovered two fundamentally distinct senescent states.
1. Transient inflammatory senescence
Observed in:
🔵 Endothelial cells
🟢 Brain macrophages
These cells appeared during mid-gestation and disappeared after barrier formation.
Their SASP programs were highly inflammatory, producing:
• IL-1β
• IL-6
• TNF
• CCL chemokines
• angiogenic factors.
These signals coordinated vascular growth and remodeling.
2. Persistent non-inflammatory senescence
Observed in:
🟡 Choroid plexus epithelial cells.
Remarkably, these cells retained senescence markers:
✅ At postnatal day 21
✅ At 12 months of age
while maintaining normal physiological function.
Unlike classical aging-associated senescence, ChPe cells lacked inflammatory SASP features and instead expressed genes supporting:
🧬 Barrier integrity
🧬 CSF production
🧬 Ciliogenesis
🧬 Ion transport
🧬 Tissue homeostasis.
This may represent the first example of a lifelong physiological senescent state in a normal somatic tissue.
Senescent endothelial cells guide vascular patterning
One of the most surprising findings was that senescence strongly overlapped with:
🌱 endothelial tip cells.
Tip cells lead angiogenic sprouts during vessel formation.
Approximately:
📈 39% of endothelial tip cells were p21⁺
compared with only ~11% of non-tip endothelial cells.
These cells expressed:
• DLL4
• KDR
• CXCR4
• angiogenic signaling programs.
The data suggest that developmental senescence helps coordinate controlled angiogenesis rather than simply terminating proliferation.
Macrophages and endothelial cells form a senescence circuit
Single-cell analyses identified extensive communication between:
🟢 Senescent macrophages
and
🔵 Senescent endothelial cells.
Key signaling axes included:
Macrophage IL-1β
→ endothelial IL1R1
and
Endothelial CSF1 / IL34
→ macrophage CSF1R.
Senescent macrophages preferentially localized near senescent blood vessels during embryonic development, suggesting a coordinated vascular remodeling niche.
Eliminating senescent cells disrupts brain development
The strongest evidence came from genetic ablation experiments.
Removing p21⁺ cells during embryogenesis caused:
☠ Embryonic lethality
🩸 Cerebral hemorrhage
🧠 Ventricular collapse
📉 Impaired CSF production
📉 Choroid plexus hypoplasia
📉 Abnormal vascular architecture.
More than 75% of embryos developed hemorrhage after p21⁺ cell depletion.
Blood vessels became:
⬆ Hyperbranched
⬆ Disorganized
⬆ Endothelial proliferation
⬇ Network complexity.
The findings indicate that senescent cells actively restrain and sculpt vascular development.
Choroid plexus senescence supports CSF homeostasis
Ablation of senescent ChPe cells produced dramatic defects.
Investigators observed:
⬇ Choroid plexus size
⬇ AQP1 expression
⬇ Endothelial support
⬇ Collagen IV deposition
⬇ Ventricular fluid volume
⬇ CSF production.
MRI analysis showed:
📉 ~45% reduction in lateral ventricular fluid volume.
Barrier integrity was also compromised, with increased erythrocyte leakage into ventricular spaces.
A new model of senescence biology
The prevailing model states:
Senescence
→ aging
→ inflammation
→ pathology.
This work suggests a more nuanced framework:
Developmental senescence
→ tissue patterning
→ barrier formation
→ physiological homeostasis.
Only later, when dysregulated or chronically activated, might senescence contribute to disease.
Why this matters
The study fundamentally changes how we think about senescence.
Instead of being solely an aging-associated damage response, senescence appears to be an ancient developmental program that can be deployed in multiple ways:
🔹 Transient inflammatory senescence to coordinate angiogenesis
🔹 Persistent non-inflammatory senescence to support lifelong epithelial function.
The findings also raise important questions for senolytic therapies.
If some senescent cells are required for tissue integrity and barrier function, indiscriminate elimination of all senescent cells may not always be beneficial.
Understanding the difference between:
✅ beneficial physiological senescence
and
❌ pathological aging-associated senescence
may become one of the central challenges of geroscience over the next decade.
Reference
Watson LA, Adelsheim Z, Carter MJ, et al.
Persistent and transient senescent cells contribute to brain-barrier development.
Cell (2026)
DOI: 10.1016/j.cell.2026.05.022
#Senescence #Aging #BrainDevelopment #BloodBrainBarrier #ChoroidPlexus #CSF #Neurobiology #DevelopmentalBiology #CellJournal #Geroscience

1
3
16
348

25 Dec 2025
✨#WCH publishes in @CellPressNews, revealing TFAP2β as a novel therapeutic target for esophageal squamous cell carcinoma—and A6, the world’s first phase-separation–targeting small molecule. A true 0→1 leap with global impact 🌍🧬#CellJournal #CancerResearch

3
120

9 Dec 2025
Huge congratulations to Alex Kwan, PhD, 2022 One Mind @COMPASSPathway Rising Star Awardee, on the publication of his groundbreaking new paper in Cell (@CellPressNews), one of the world’s most prestigious scientific journals.
This publication marks an exciting milestone for Dr. Kwan’s research, which was supported in part by his 2022 One Mind Rising Star Award. That support helped his team advance a bold line of inquiry into how psychedelics like psilocybin influence large-scale brain circuit plasticity. This work is now shedding new light on how the brain adapts, heals, and responds to treatment.
Dr. Kwan’s team has uncovered remarkable insights into how psychedelics spark widespread rewiring across brain circuits. This kind of circuit-level understanding is essential for opening new therapeutic possibilities for depression and other serious mental health conditions.
Dr. Kwan's work reflects the mission of the One Mind Rising Star Awards, which is to empower early-career scientists who are asking big questions and pushing the boundaries of what is possible in brain research. Their discoveries not only advance science but also have the potential to reshape lives.
Congratulations, Alex. We are excited to see how this research continues to evolve.
⭐️ Learn more about the One Mind Rising Star Awards: onemind.org/what-we-do/one-m…
📰 Read the paper: cell.com/cell/fulltext/S0092…
#Neuroscience #Psychedelics #Research #CellJournal #BrainScience #Innovation #OneMind #CellPress
1
3
11
1,863

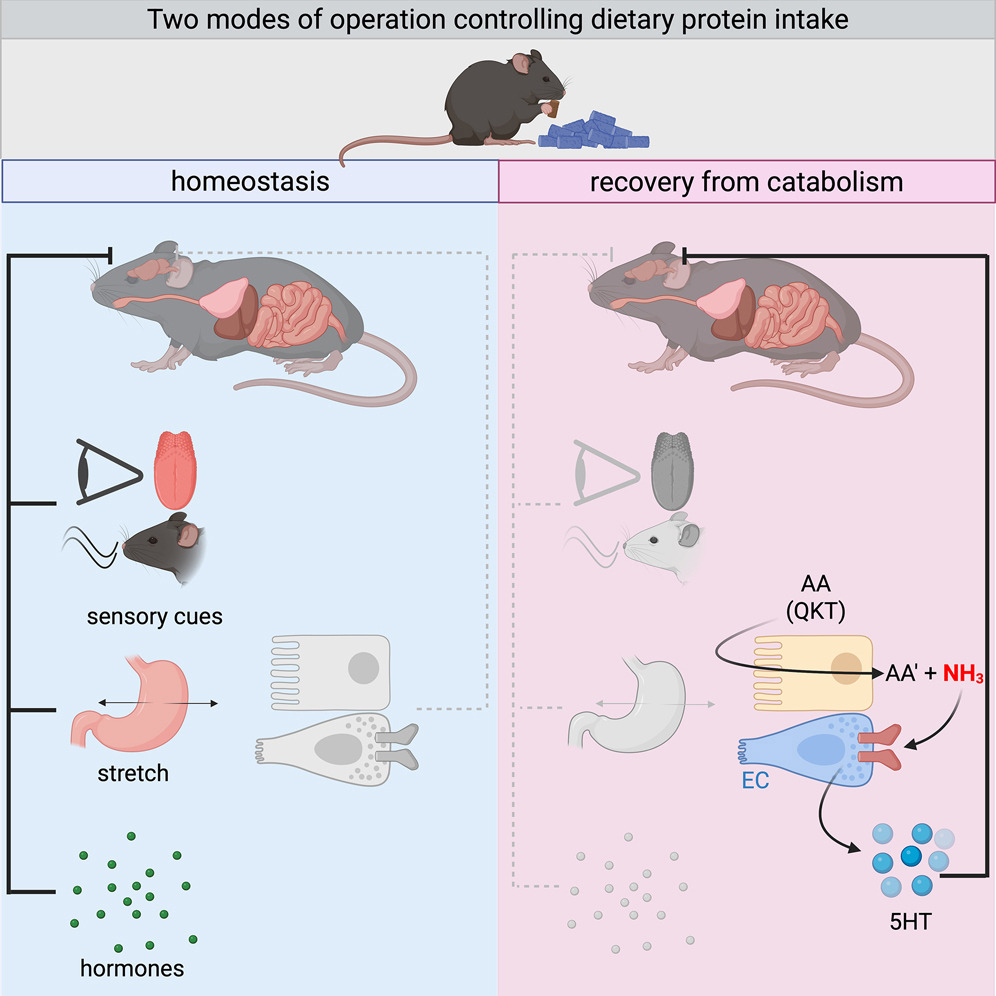
🥩 Why Your Body Rejects Meat During Recovery
When you're healing, this instinct isn't random — it's biochemistry.
🧠 Explore more breakthrough biology on PubMed.ai → pubmed-ai-newsletter.beehiiv…
New Cell research uncovers a gut–brain circuit that explains protein aversion during illness:
Three amino acids — glutamine, lysine, threonine — ramp up ammonia production in the gut.
Enterochromaffin cells sense this via TRPA1, signal the brainstem through serotonin, and trigger anorexia.
It’s a built-in safety mechanism: your body limiting ammonia toxicity during catabolic stress.
#MedWeekly #PubMedAI #GutBrainAxis #CellJournal #Immunometabolism #AminoAcids #ScienceTwitter #MedTwitter #BiomedicalResearch #HealthTech
2
7
266

🧬 〈Deep Dive〉How exercise rewires gut metabolism to enhance cancer immunity.
A Cell 2025 study (PMID 40639377) reveals that
exercise activates the one-carbon metabolism of gut microbiota,
increasing production of formate, a metabolite that:
•boosts CD8 T-cell metabolism and cytokine release,
•suppresses tumor growth, and
•enhances checkpoint-inhibitor efficacy.
Eliminating gut bacteria abolished this benefit, proving it’s microbiota-dependent.
💡 Exercise may thus act as a natural adjuvant for cancer immunotherapy.
📖 Cell 2025; PMID 40639377
🔗 pubmed.ncbi.nlm.nih.gov/4063…
⸻
#Microbiome #CancerImmunity #ExerciseScience #Formate #Immunotherapy #MetabolicImmunology #CellJournal
4
133
🏃♂️ Exercise boosts cancer immunity!
A Cell 2025 study found that exercise makes gut microbes produce
formate, a metabolite that strengthens CD8 T-cell antitumor immunity
and even enhances immune-checkpoint inhibitor efficacy.
In short:
Exercise doesn’t just build muscle — it empowers your immune system to fight cancer.
📖 Phelps CM et al. Cell. 2025; PMID 40639377
🔗 pubmed.ncbi.nlm.nih.gov/4063…
⸻
#Exercise #Microbiome #CancerImmunity #Immunotherapy #Formate #LongevityScience #CellJournal
3
106

29 Aug 2025
New data published in Cell show that Decipher® Prostate Genomic Classifier predicts which patients with metastatic cancer are likely to benefit from treatment intensification with docetaxel chemotherapy, as well as those who are not and may therefore avoid unnecessary toxicity.
These findings, from the randomized, prospective, Phase 3 STAMPEDE trial, are the first to be published showing that a gene expression test can help clinicians better personalize chemotherapy decisions for patients with metastatic prostate cancer.
Read more: investor.veracyte.com/news-r…
#DecipherUrologicCancers #ProstateCancer #STAMPEDE #CellJournal #PrecisionMedicine #biotech
Disclaimer: veracyte.com/cautionary-note…

3
6
331

27 Aug 2025
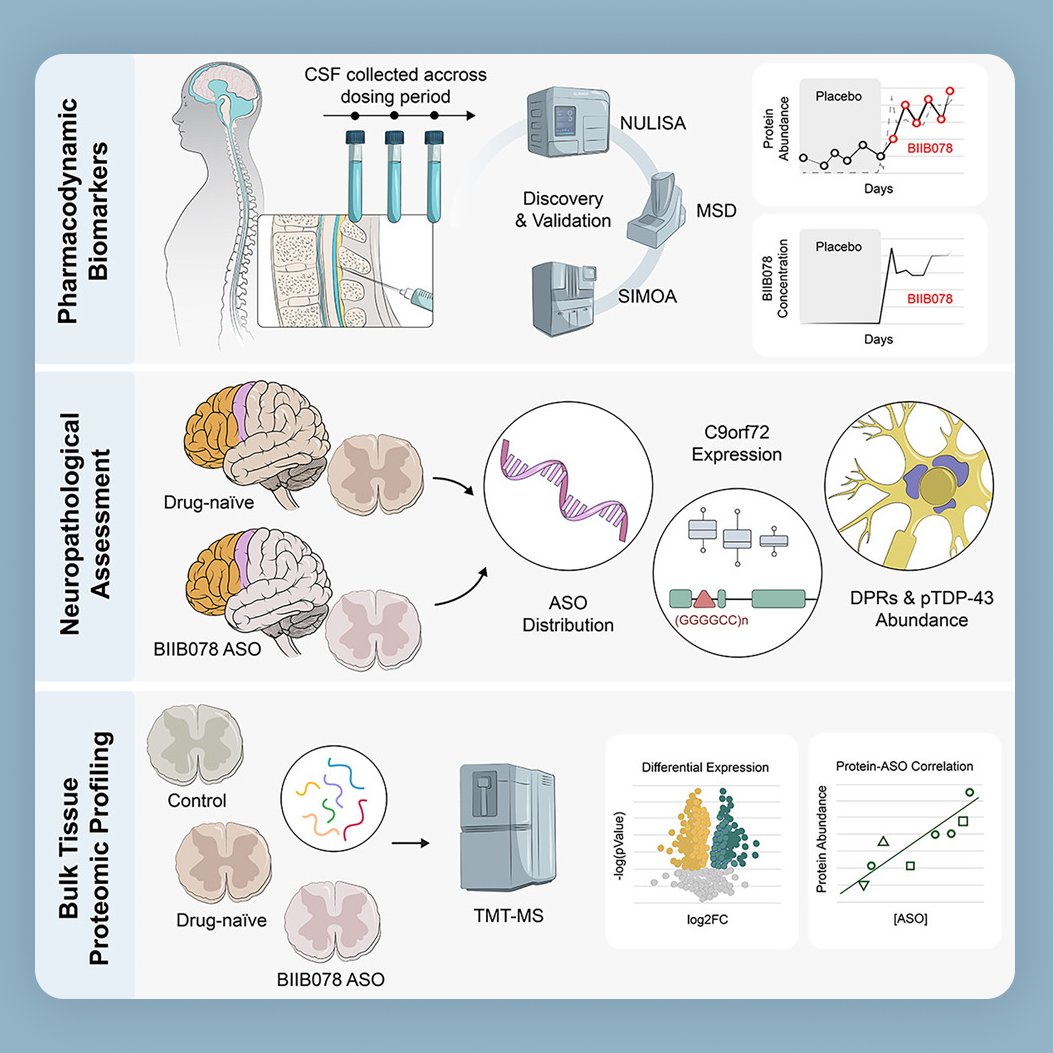
👀 Spotted in @CellCellPress ! Excited to see one of our graphical abstracts featured in this new paper on C9orf72-associated ALS (c9ALS).
🧠 The study looks at an antisense oligonucleotide (ASO) therapy designed to reduce toxic RNA and proteins linked to this form of ALS. The results show the treatment reached the brain and spinal cord, lowered some of the harmful markers, but also highlight how challenging it is to fully reverse disease pathways, pointing to the need for better biomarkers in the future.
Always a pleasure turning such impactful science into a clear visual story. Congratulations to the authors on this important work!
#JonathanGlass #ZacharyMcEachin
👉 cell.com/cell/fulltext/S0092…
---
👩🏻🎨 Hi, I’m Emma Vidal, founder and illustrator at @DrawImpacts . We help scientists, labs, and institutions bring their research to life with visuals that are clear, compelling, and memorable, from journal covers and graphical abstracts to lab logos and websites. drawimpacts.com
#ALS #Neurodegeneration #GraphicalAbstract #DataViz #ScientificIllustration #ScienceCommunication #CellJournal #DrawImpacts
1
13
1,384

1 Aug 2025
Tomato is the mother of potato?
Chinese scientists, in collaboration with Canadian and British counterparts, revealed that the potato originated from an ancient hybridization event between the tomato plant and a potato-like plant about 9 million years ago. Published in the latest issue of the Cell journal, these findings provide a groundbreaking theoretical perspective for the genetic breeding of potatoes.Source:People's Daily,China #Tomoto #potato #Canadian #British #counterparts #CellJournal

2
17
176

26 Apr 2024
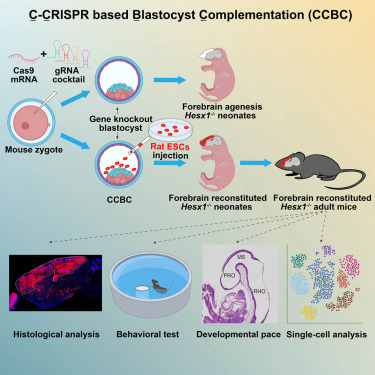
Breaking in Cell: Scientists create "hybrid brains" using different species. #Neuroscience #Innovation #CellJournal @Innov_Medicine
25 Apr 2024

Generation of rat forebrain tissues in mice: Cell cell.com/cell/abstract/S0092…
3
6
4,488

2 Apr 2024
'Hallmarks of Cancer' paper gets an upgrade to v 3.0 in this issue of Cell - @DrHBurstein
@CellCellPress
#CancerCell #CellJournal #HallmarksOfCancer #OncoDaily #Oncology
oncodaily.com/43487.html
1
13
464

16 Oct 2023
Share another interesting study on tomato breeding: "Rewiring of the Fruit Metabolome in Tomato Breeding," published in Cell, using Metware Widely-Targeted #Metabolomics assay. Discover more at: metwarebio.com/metabolomics-…
#TomatoBreeding #Research #CellJournal #genetics
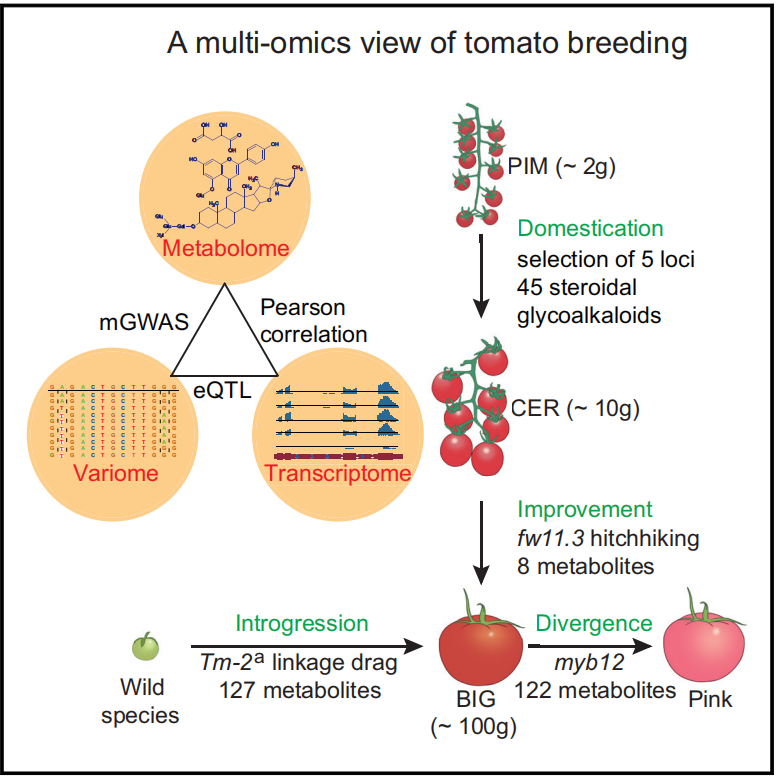
ALT A multi-omics view of tomato breeding Zhu et al., 2018, Cell
15 Oct 2023

Jose Blanca @jblanca42 closes the physiology session at the #SOL2023_Montreal talking about the evolution and history of the tomato fruit shape. They analyze the G2P-SOL collection with SPET. First cultivated tomato in Europe were flat and highly ribbed (1/3).

2
8
313

7 Aug 2023
Thrilled to be part of this study published in Cell! Huge congratulations to Yuanyuan Liu, Dylan Dodd, and the entire team of authors! Read the full article here - cell.com/cell/pdf/S0092-8674… #CellJournal #GutMicrobe
1
15
2,508